Construction and characterization of recombinant epitope protein
We screened some B cell epitopes and T cell epitopes of the GP5 and M protein of PRRSV identified in other studies(16–18), according to the location and size of the epitopes, some of the epitopes have been selected as candidate epitopes and all selected epitopes were listed in Table 1. B cell epitopes and T cell epitopes of P46 and P65 protein of Mhp were predicted with Bepipred Linear Epitope Prediction and ProPred-I. According to the position and length of the epitopes, several candidate epitopes were screened out for the construction of recombinant protein (Table 2).
Table 1
Candidate epitopes of GP5 and M protein of PRRSV.
|
Type of epitope
|
Protein
|
Sequence of amino acid
|
|
B cell
|
GP5
|
SSHIQLIYNLTLCELNG
|
|
B cell
|
GP5
|
VEKGGKVEVEGHLIDLKRV
|
|
T cell
|
GP5
|
LAALICFVIRLAKNC
|
|
T cell
|
GP5
|
RLYRWRSPV
|
|
B cell
|
M
|
FVVRRPGSTTVNGTLVPGLKSLVLGGR
|
|
B cell
|
M
|
AVKQGVVNLVKYAK
|
|
T cell
|
M
|
CNDSTAPQKVLLAFS
|
|
T cell
|
M
|
FGYMTFVHFESTNRV
|
Table 2
Candidate epitopes of P46 and P65 protein of Mhp.
|
Type of epitope
|
Protein
|
Sequence of amino acid
|
|
B cell
|
P46
|
ANLSPAPKGFIIAPENGSGVGTA
|
|
B cell
|
P46
|
KYDNQTYKVQGK
|
|
T cell
|
P46
|
KKFLYSSAIYA
|
|
T cell
|
P46
|
DLSPEGENAVYV
|
|
B cell
|
P65
|
DQEEKDDSNAEELKNTTNFDDFDENKPTYS
|
|
B cell
|
P65
|
STFDTDQEAAIKDDKRT
|
|
B cell
|
P65
|
NKVKDYAR
|
|
T cell
|
P65
|
GYKKIAHQLLLKLTLDQEEK
|
|
T cell
|
P65
|
VREIVSLPIFDNFDFRELIPVKNPFV
|
Candidate epitopes were concatenated together to construct the recombinant epitope protein rEP-PM, the linker KK had been used between T-T and T-B epitopes, GGGGS had been used to connect B-B epitopes (Fig. 1A). The hydrophilicity plot, antigenic index, surface probability plot and flexible regions were analyzed by using the Kyte-Doolittle, Jameson-Wolf, and Emini algorithms, respectively, with the aid of DNASTAR package (Fig. 1B). The results indicate that rEP-PM has good hydrophilicity, strong antigenicity and high surface accessibility, and each epitope is independent and complete.
The secondary structure of rEP-PM was predicted by SOPMA server, the result showed that rEP-PM consists of 21.51% alpha helix (h), 27.91% extended strand (e), 12.5% beta turn (t) and 38.08% random coil (c) secondary structural elements (Fig. 1C).
Expression and purification of the rEP-PM
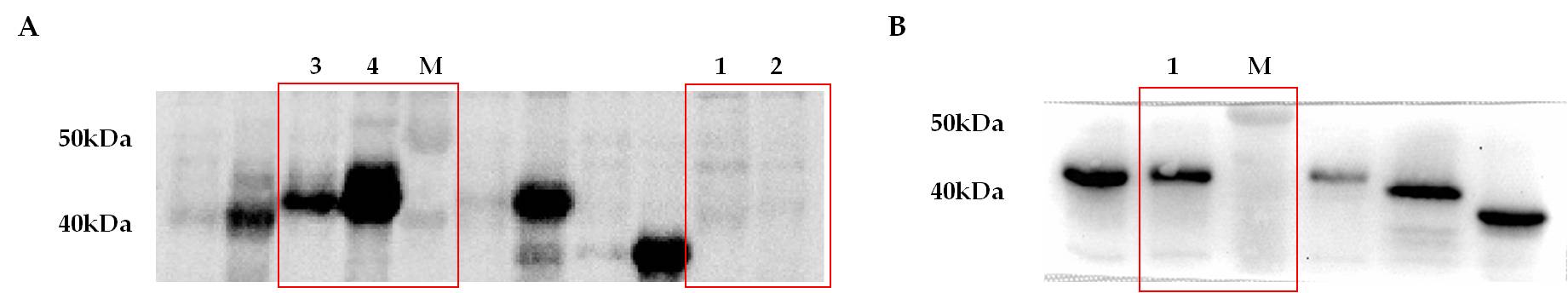
DNA sequence encoding rEP-PM was synthesized and cloned into the plasmid pET-28a, the supernatant and cell lysate were derived from E.coli BL21 transformed with pET-28a-rEP-PM and analyzed by western blot. A single protein band with a molecular mass of approximately 41kDa could be observed by anti-His monoclonal antibodies in the supernatant and cell lysate, no specific protein band has been detected in control group (Fig. 2A). This result showed that the rEP-PM was partially expressed in soluble form. Then the soluble rEP-PM in supernatant of E.coli BL21 has been purified with nickel affinity chromatography column (Fig. 2B). All these results demonstrated that the rEP-PM was successfully expressed and purified.
rEP-PM could induce the production of high levels of antibodies in mice
To investigate whether rEP-PM could induce specific antibodies, four-weeks-old female BALB/c mice were divided into four groups of twelve mice in each group, mice were immunized as described in Materials and methods. Serum samples were taken from the twelve mice in each group at day 7, 14, 28 and 56 after the third immunization, the levels of antibodies in the serum of mice were assessed with indirect ELISA, the antibody titers of mice immunized with rEP-PM were higher than 12800 at day 7 (Fig. 3A), 14 (Fig. 3B), 28 (Fig. 3C) and 56 (Fig. 3D) after the third immunization. These results indicate that mice immunized with rEP-PM could induce high levels of antibodies, which maintained at high levels for a long time (8 weeks) after the last immunization.
rEP-PM has good immunoreactivity
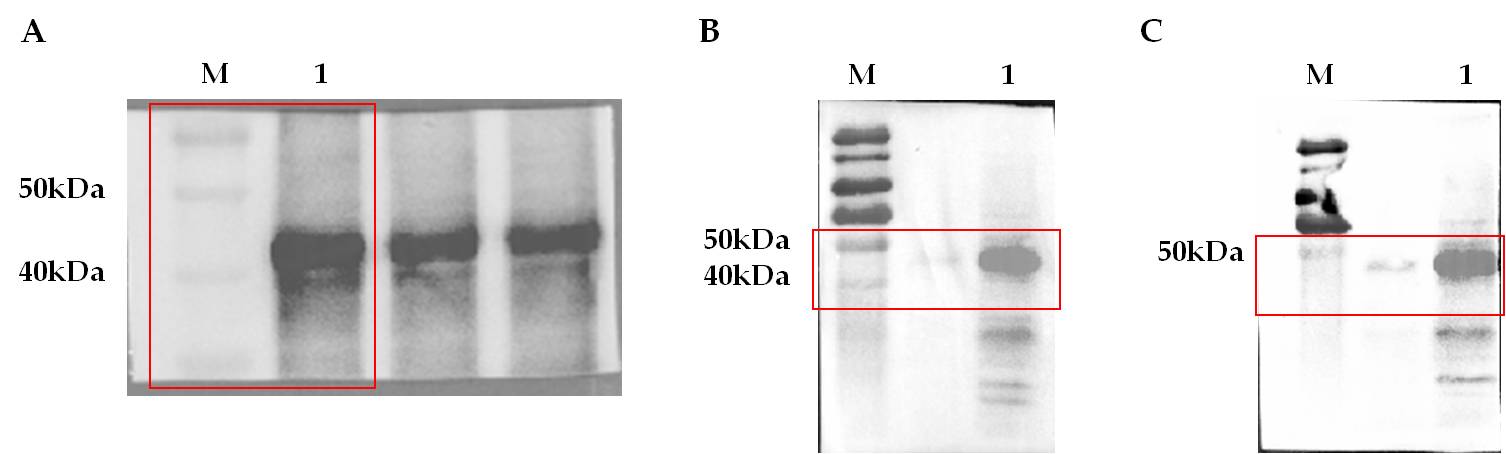
The specific immunoreactivity of rEP-PM in mice was further confirmed by western blot. 10 µg of purified rEP-PM was subjected to polyacrylamide gel electrophoresis, the serum of mice immunized with rEP-PM protein was used as the primary antibody and the goat anti-mouse IgG as the secondary antibody. The result indicate that rEP-PM could react specifically with antibodies induced by rEP-PM (Fig. 4A).
To further explore whether rEP-PM could react with anti-PRRSV antibodies and anti-Mhp antibodies, 10 µg of purified rEP-PM was subjected to polyacrylamide gel electrophoresis, the serum of mice immunized with PRRSV attenuated vaccine (containing anti-PRRSV antibodies) and Mhp-168 attenuated vaccine (containing anti-Mhp antibodies) were used as primary antibodies, respectively, and the goat anti-mouse IgG was used as the secondary antibody. As shown in Fig. 4B and 4C, the recombinant protein rEP-PM could react specifically both with anti-PRRSV (Fig. 4B) and anti-Mhp (Fig. 4C) antibodies. These data suggest that rEP-PM has good immunoreactivity with anti- rEP-PM, anti-PRRSV and anti-Mhp antibodies.
rEP-PM could induce Th1 and Th2 cellar immune response in mice
To investigate whether rEP-PM could induce Th1 and Th2 cellular response, the concentration of IL-2 (which induced by Th1 cellular response) and IL-10 (which induced by Th2 cellular response) in the medium supernatant of splenic lymphocytes were detected by ELISA. As shown in Fig. 5, compared with the negative control group, both the level of IL-2 and IL-10 in the medium supernatant of splenic lymphocyte of the immunized groups was significantly higher than the negative control group. These results suggest that immunized with rEP-PM could induce both Th1 and Th2 cellular immune response in mice.
{kind=link}
{kind=link}