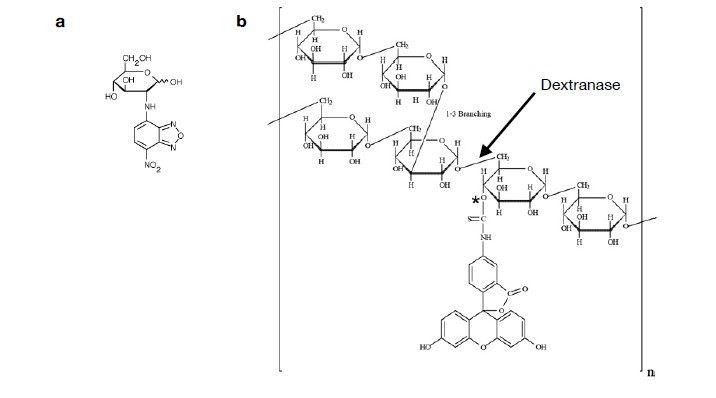
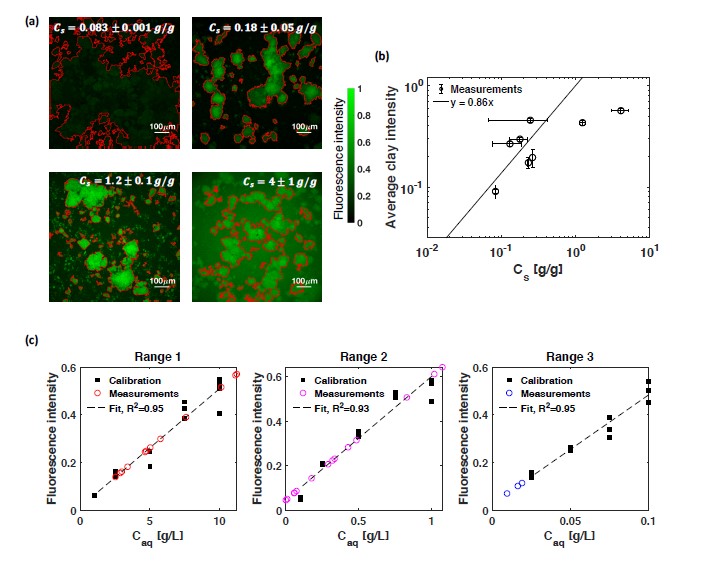
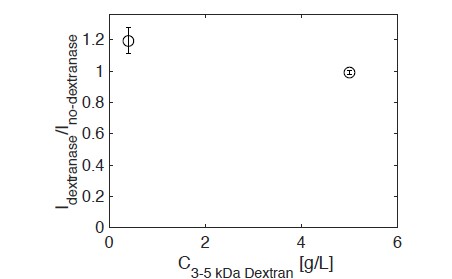
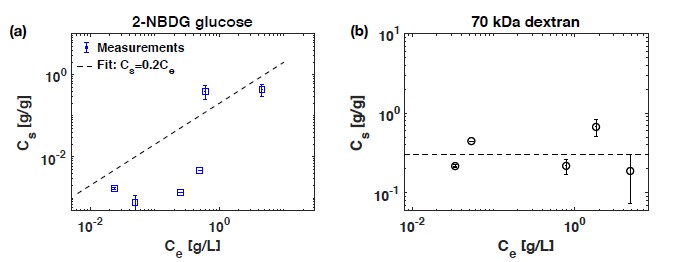
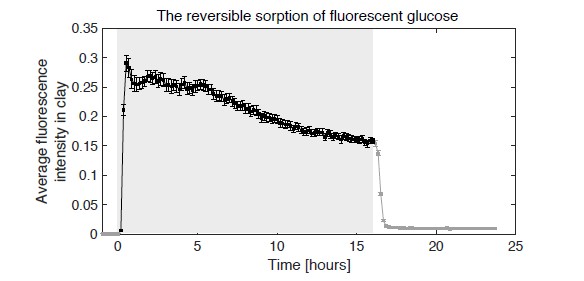
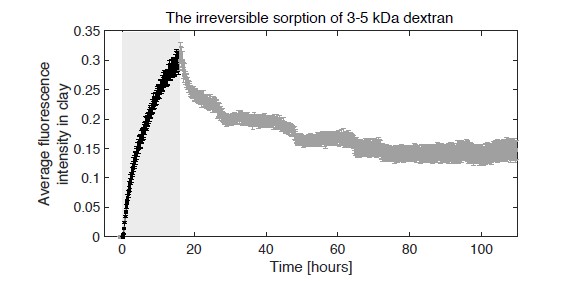
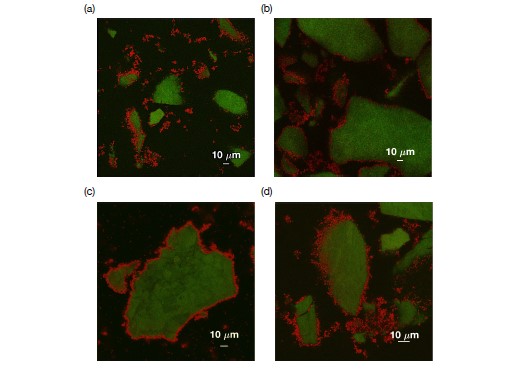
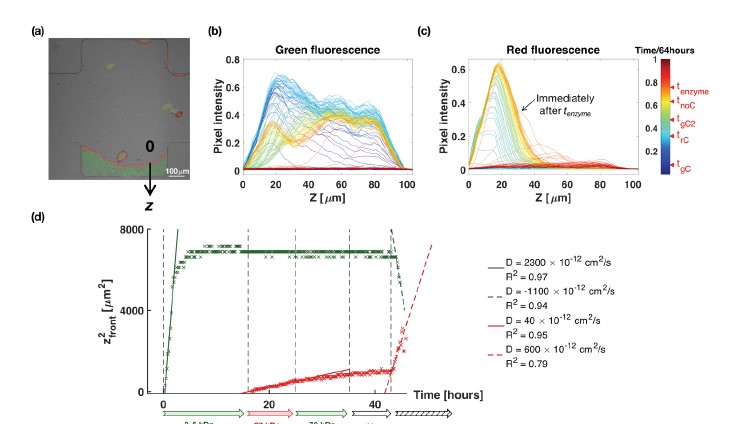
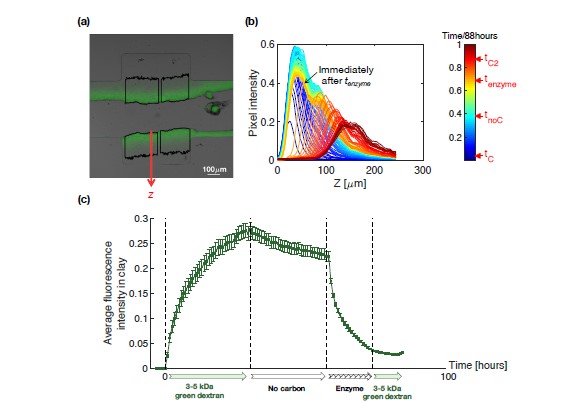
Soil absorbs about 20% of anthropogenic CO2 emissions annually, and clay is the key carbon-capture material. Although sorption to clay is widely assumed to strongly retard the microbial decomposition of soil organic matter, enhanced degradation of clay-associated organic carbon has been observed under certain conditions. The conditions in which clay inhibits microbial decomposition remain uncertain because the mechanisms of clay-organic carbon interactions are not fully understood. Here we reveal the spatiotemporal dynamics of carbon sorption and release within clay aggregates and the role of enzymatic decomposition by directly imaging a transparent smectite clay on a microfluidic chip. We demonstrate that clay-carbon protection is due to the quasi-irreversible sorption of high molecular-weight sugars within clay aggregates and the exclusion of bacteria from these aggregates. We show that this physically-protected carbon can be enzymatically broken down into fragments that are released into solution. Further, we suggest improvements relevant to soil carbon models.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}