NALCN, a sodium leak channel mainly expressed in the central nervous systems, is responsible for the resting Na+ permeability that controls neuronal excitability. Dysfunctions of the NALCN channelosome, NALCN with several auxiliary subunits, are associated with a variety of human diseases. Here, we reported the cryo-EM structure of human NALCN in complex with FAM155A, at an overall resolution of 3.1 angstrom. FAM155A forms extensive interactions with the extracellular loops of NALCN that help stabilize NALCN in the membrane. A Na+ ion-binding site, reminiscent of a Ca2+ binding site in Cav channels, is identified in the unique EEKE selectivity filter. Despite its ‘leaky’ nature, the intracellular gate is sealed by S6I, II-III linker and III-IV linker. Our study establishes the molecular basis of Na+ permeation and voltage sensitivity, and provides important clues to the mechanistic understanding of NALCN regulation and NALCN channelosome-related diseases.
Article
Structure of human sodium leak channel NALCN in complex with FAM155A
https://doi.org/10.21203/rs.3.rs-56923/v1
This work is licensed under a CC BY 4.0 License
Journal Publication
published 16 Nov, 2020
Version 1
posted
You are reading this latest preprint version
NALCN
Na+ permeation
Na+ voltage
regulation
channelosome-related diseases
Membrane potential across the cell membrane is essential for signal transduction in excitable cells such as neurons and muscle cells 1. In neurons, the resting membrane potential (RMP) is approximately − 70 mV, considerably depolarized as compared to the equilibrium potential of K+ of -90 mV. The depolarization is mostly attributable to the resting Na+ permeability, which is important for the regulation of neuronal excitability 2,3. The newly identified ion channel NALCN (sodium leak channel, non-selective protein) is primarily expressed in the central nervous system 4. It is mainly responsible for the neuronal tetrodotoxin (TTX) -resistant Na+ leak conductance and plays a key role in regulating RMP and controlling neuronal excitability 5.
NALCN consists of a single polypeptide chain of 24 transmembrane helices (TM) that form four homologous functional repeats connected by intracellular linkers. The topology of NALCN is similar to that of voltage-gated sodium (Nav) channels and voltage-gated calcium (Cav) channels (4 × 6 TM). Each functional repeat of NALCN contains six TMs (S1-S6), with S1-S4 corresponding to the voltage-sensing domain (VSD) in Nav/Cav. S5, S6 and intervening segments, including the pore helices and selectivity filter (SF) from all four repeats, constitute the ion conducting pore. NALCN shares less than 20% identity with any member of Nav and Cav channels, and it has been classified as a member of a new subclass of the 4 × 6 TM ion channel family. Compared to Nav and Cav channels, NALCN contains fewer positively charged residues (arginine or lysine) on S4 segments. Additionally, NALCN has a unique ion selectivity filter (SF) with EEKE residues in the four repeats, which is different from SFs of Nav channels (DEKA) and Cav channels (EEEE/EEDD) 4.
NALCN is associated with several auxiliary subunits, including UNC80, UNC79 and FAM155A. Together they form a large protein complex termed the NALCN channelosome. Notably, these auxiliary subunits are unique to NALCN and share no sequence homology to the auxiliary subunits of any other channel. Both UNC80 and UNC79 are large proteins that contribute to the neuronal localization and stabilization of NALCN in C. elegans and D. melanogaster, although their own functional domains and subcellular localization are largely unclear 6–8. UNC80 interacts with NALCN directly and acts as a scaffold protein for UNC79 9,10. FAM155A, also named NLF-1 in C. elegans, was reported to be an endoplasmic reticulum (ER) resident protein that acts as a chaperone to facilitate the folding and promotes axon delivery of NALCN 11. In humans, FAM155B, a FAM155A homolog, can functionally substitute for FAM155A, and is likely a component of the NALCN channelosome. In addition, NALCN channelosome activity is regulated by the Src family of tyrosine kinases (SFKs) and several GPCRs such as the M3 muscarinic receptor (M3R) through direct interactions 12. The NALCN channel is also activated by neuropeptides, including substance P and neurotensin, through the SFK-dependent pathway in mouse hippocampal neurons 13. How NALCN interacts with and is regulated by its auxiliary subunits remains largely unclear.
NALCN is evolutionally conserved in both invertebrate and vertebrate species. For example, NALCN homolog have been identified in invertebrate species such as snails and C. elegans, in which Nav homologues were not found 4,14. This indicates that NALCN evolved earlier than Nav channels across species. NALCN is highly conserved in mammals, with the sequence identity of more than 96% among human, mouse, rat, rabbit and bovine. Human NALCN also shares at least 44% identity and 56% similarity with its homologs α1U in D. melanogaster and NCA-1/2 in C. elegans.
Except for the role in regulating neuronal excitability, NALCN is also important in many fundamental physiological processes, such as motor function, pain sensitivity and circadian rhythm. For example, NALCN mutant mice die within 24 hours after birth due to disrupted respiratory rhythm 5. Overexpression of NALCN in highveld mole-rat leads to ablating detection of painful substances by nociceptors 15. Reduced NALCN expression in Drosophila leads to changes in behavioral circadian rhythms and sensitivity to anesthetics 16,17. In humans, NALCN variants are linked to a variety of diseases, including congenital contractures of limbs and face, hypotonia and developmental delay (CLIFAHDD)18–20, psychomotor retardation and characteristic facies (IHPRF) 21–23, infantile neuroaxonal dystrophy (INAD) 24, cervical dystonia, schizophrenia and bipolar disorder 25–27. There are also many diseases related to the dysfunction of the auxiliary subunits of NALCN 28,29.
Despite its physiological and pathological importance, the biophysical properties of the voltage sensitivity and ion selectivity of NALCN are still under debate. Electrophysiological studies on NALCN have been performed in heterologous expression systems such as HEK293 cells, X. laevis oocytes and the neuronal cell line NG108-15 5,30,31. NALCN expressed in HEK293 cells was reported to exhibit a linear current-voltage relationship, suggesting that NALCN is a voltage-independent ion channel. The same study also reported that NALCN is nonselective and permeable to Na+, K+ and Ca2+ 5. Other studies suggested that the VSDs of NALCN can exhibit a broader range of gating behaviors. Co-expression of NALCN, UNC79, UNC80, and FAM155A resulted in voltage dependent NALCN current 30,31. Finally, NALCN was also reported to be only selective for monovalent cations and to be blocked by extracellular divalent cations 30.
To better understand the functional properties and mechanisms of NALCN, we sought to determine the structure of NALCN in complex with its auxiliary subunits. Here, we report the cryo-EM structure of human NALCN in complex with FAM155A at an overall resolution of 3.1 Å. Our structure along with electrophysiology and molecular dynamic (MD) simulation data, provide unprecedented insights in understanding the structure, function and regulation of the NALCN channelosome.
Functional characterization of NALCN by electrophysiology
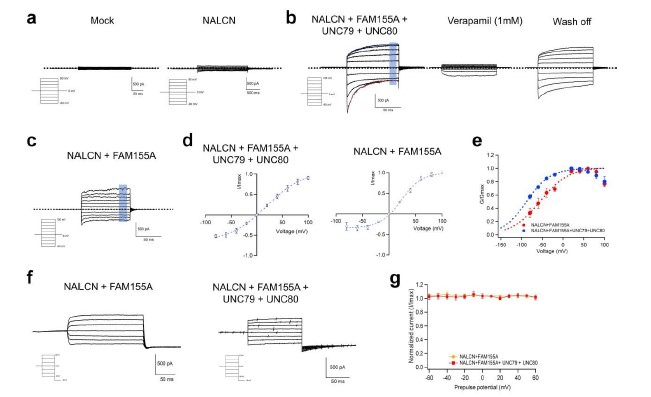
We employed patch-clamp recordings in HEK293 cells to characterize the electrophysiological properties of NALCN. The current elicited in response to voltage steps was very small and appeared to be indistinguishable from the mock when NALCN channel was expressed alone in HEK293 (Supplementary Fig. 1a). By contrast, co-expression of NALCN with UNC79, UNC80 and FAM155A dramatically enhanced the current, which was largely inhibited by 1 mM verapamil, implying that the current measured was mainly mediated by NALCN (Supplementary Fig. 1b). We also clearly recorded the kinetics of voltage sensitive NALCN current: hyperpolarization voltages elicited large inward current with inactivation, while depolarization voltages activated current with an exponential course before reaching in the steady state (Supplementary Fig. 1b). These observations indicate that the auxiliary subunits are essential for a functional NALCN (Supplementary Fig. 1a), consistent with previous studies 30,31. The electrophysiological properties of NALCN suggested that it tends to maintain the RMP in a manner reminiscent the Le Chatelier’s principle in chemical equilibria. In comparison, Nav channels always show rapid current activation and inactivation for fast initiating action potential 32, while certain Kv such as KCNQ channels show slow activation kinetics to avoid fast repolarization 33.
Structure determination of human NALCN-FAM155A complex
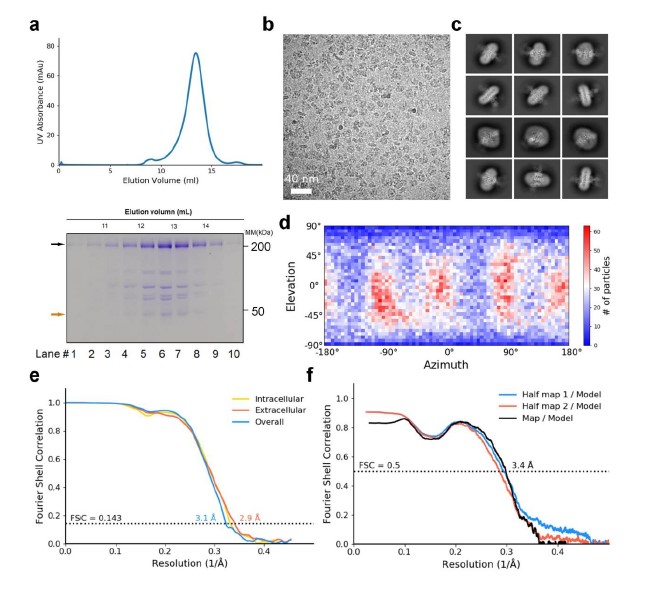
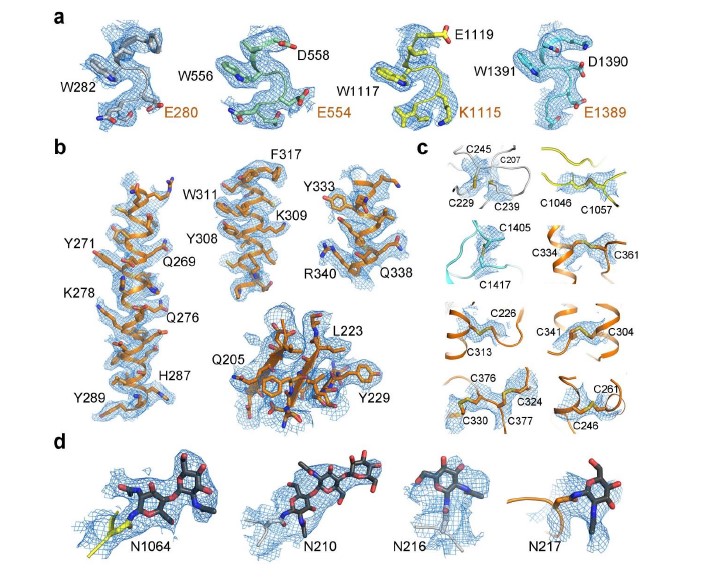
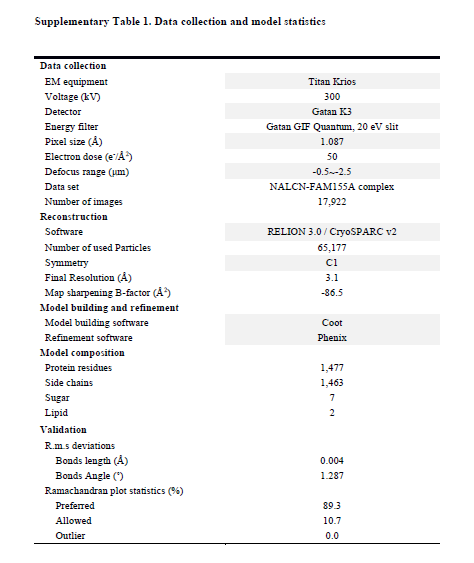
Based on our functional characterizations, we focused on the co-expression of the four components in HEK293 cells. Unfortunately, we were unable to get a stable complex of the four components. Instead, a stable and homogeneous subcomplex of NALCN and FAM155A was obtained (Supplementary Fig. 2a). The peak fractions from gel filtration purification, were pooled and concentrated to about 8.5 mg/ml for cryo-EM sample preparation. The EM images were collected on a Titan Krios electron microscope operating at 300 kV and equipped with a K3 direct detector and a Gif Quantum energy filter. After a few rounds of two-dimensional (2D) and 3D classifications, about sixty-five thousand particles were selected, which yield a final reconstruction map with an overall resolution of 3.1 Å (Fig. 1, Supplementary Table 1, Supplementary Fig. 2, 3). Local masks of the extracellular and intracellular regions were applied to further improve the local resolutions to 2.9 Å and 3.1 Å, respectively.
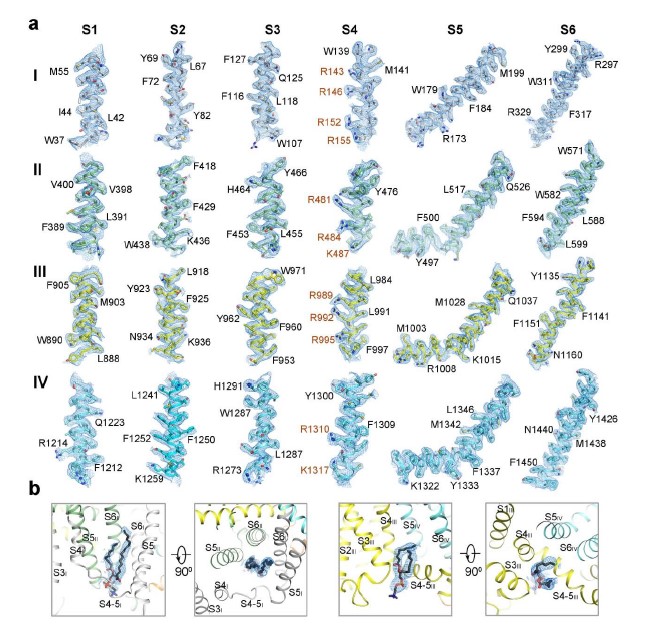
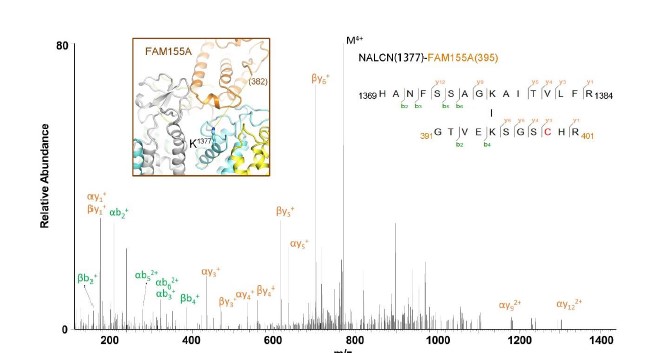
The density of the final reconstruction is of high quality with evenly distributed estimated local resolution. Most side chains were clearly resolved that allow for accurate model building (Fig. 1a, 1b, Supplementary Fig. 4, 5). The model of NALCN was built with a homology model based on Cav1.1 (PDB: 5GJV), whereas FAM155A was built de novo. In total, 1295 and 182 residues were built for NALCN and FAM155A, respectively. The complex structure was confirmed by cross-linking mass spectrometry analysis (Supplementary Fig. 6). Three glycosylation sites (Asn210, Asn216 and Asn1064) and one glycosylation site (Asn217) were identified on NALCN and FAM155A, respectively (Fig. 1c, Supplementary Fig. 5c). Four disulfide bonds in the extracellular loops (ECL) of NALCN and six in FAM155A were recognized (Fig. 1c, Supplementary Fig. 5d). The glycosylation sites and disulfide bonds in turn verified the accuracy of the structure.
The overall structure of NALCN resembles those of eukaryotic Nav and Cav channels (Fig. 2). We select human Nav1.4 (PDB: 6AGF) and rabbit Cav1.1 (PDB: 6JP5) as representatives of Nav and Cav channels for comparisons with NALCN 34,35. The structures of human Nav1.7 (PDB: 6J8I), rat Nav1.5 (6UZ3) and NavPaS (6A95) from American cockroach were also picked for specific discussions 36-38. The structures of NALCN can be superimposed to the α subunit of Nav1.4 and α1 subunit Cav1.1 with root-mean-square deviation (r.m.s.d) of 2.99 Å over 760 Cα atoms and 3.894 Å over 905 Cα atoms, respectively. Except for the four homologous repeats, a C-terminal domain (CTD) after repeat IV, which was observed in the structures of Cav1.1 and NavPaS but not in human Nav channels, was also clearly resolved in the structure of NALCN (Fig. 1c). NALCN has no N-terminal domain (NTD) before repeat I, which was always observed in human Nav channels.
NALCN has several unique features that are distinct from both Nav and Cav channels. In particular, two lipids, lipid1 near repeat I and lipid2 near repeat III, were found to bind NALCN at the inner membrane side (Fig. 1c). Lipid2 resides in a cavity surrounded by S3, S4 and S4-5 of repeat III, while lipid1 occupies the cavity surrounded by S4I, S5I, S6I, S5II, S6II and S4-5I (Fig. 2c, Supplementary Fig. 4b). Notably, lipid1 sits in a position that occupied by S4-5 linker in Nav/Cav structures (Fig. 2c). This may have caused the conformational change of the S4-5I linker from a helix to a loop in NALCN. In addition, a short helix designated as the II-III linker, that was connected to S6II by a short loop was firstly observed in the structure of NALCN (Fig. 1c).
Specific interactions between NALCN and FAM155A
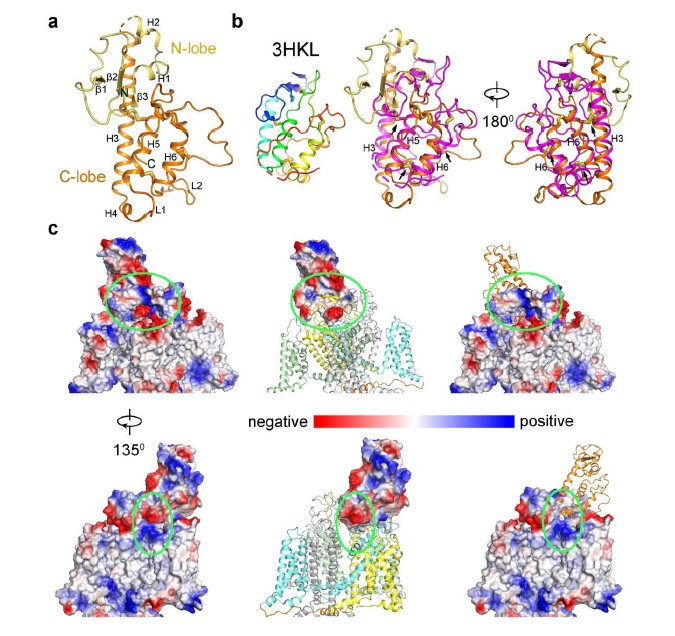
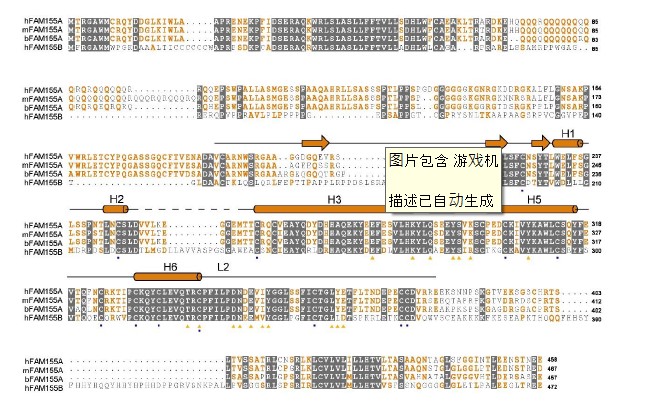
Human FAM155A contains 456 amino acids and residue 192-382 were resolved in our structure except for a short loop from residue 250 to 258 (Fig. 3a). The N-terminal and the C-terminal regions of FAM155A, including two predicted TM helices, were not resolved in current structure, probably because of their intrinsic flexibility. The resolved structure of FAM155A contains two lobes, designated as N-lobe and C-lobe, stabilized by six disulfide bonds. The N-lobe consists of three short β strands and two short α helices (H1-H2), while the C-lobe contains four α helices (H3-H6) (Fig. 3b). The overall structure of FAM155A is different from any auxiliary subunit in Nav or Cav channels (Fig. 2b, Supplementary Fig. 7a). A search of the protein data bank using the DALI server 39 revealed that the C-lobe of FAM155A contains a fold (similarity Z-score 4.3) similar to the structure of the frizzled-like cysteine-rich domain (CRD) of receptor tyrosine kinase MuSK (PDB: 3HKL), which had 84 Cα atoms aligned to FAM155A with a r.m.s.d. of 2.8 Å (Supplementary Fig. 7b). Superimposition of the CRD domain of MuSK on FAM155A shows that H3, H5 and H6 align quite well. Moreover, three disulfide bonds are structurally conserved between the two structures (Supplementary Fig. 7b).
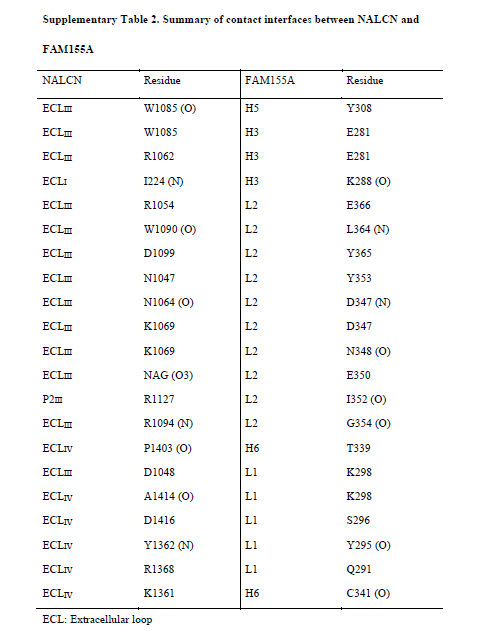
Only the C-lobe of FAM155A is involved in the interaction with NALCN. FAM155A sits above the pore domain of repeat IV of NALCN and form extensive interactions with the ECLI, ECLIII and ECLIV of NALCN (Fig. 1c, 3c, Supplementary Table 2). The loop in FAM155A connecting H4 and H5 and a hairpin loop after H6 that is involved in binding to NALCN are designated L1 and L2, respectively (Fig. 3b). Notably, L2 of FAM155A inserts deeply into a dome formed by ECLIII of NALCN. The H3, H5, H6 helices and L2 of FAM155A resemble the palm and thumb of a hand, respectively, holding ECLIII of NALCN tightly (Fig. 3c). Detailed analysis of the interaction interfaces revealed extensive salt bridges and hydrogen bonds between NALCN and FAM155A (Fig. 3d, Supplementary Table 2). Unexpectedly, the sugar moiety linked to Asn1064 on ECLIII, also contributes to the interaction, by forming a hydrogen bond with E350 on FAM155A (Fig. 3d). Electrostatic surface analysis revealed that two surfaces between NALCN and FAM155A are electrically complementary, ensuring a favorable interaction stability (Supplementary Fig. 7c). Sequence alignment among NALCN and Nav/Cav channels revealed that the interface residues in NALCN, including Asn1064, are not conserved among other Nav/Cav channels, explaining the high binding specificity of FAM155A towards NALCN, but not other channels (Supplementary Fig. 8). The interface between FAM155A and NALCN is very different from those between the ion conducting α subunit of Nav and Cav channels and their auxiliary subunits (Fig. 2b). The key residues in the interaction interface are highly conserved among NALCN and FAM155A proteins from different species, indicating that the interaction mode between NALCN and FAM155A is evolutionarily conserved. Furthermore, the key residues mediating complex formation in FAM155A are also conserved in FAM155B, another subtype protein of the FAM155 family (Supplementary Fig. 9). Therefore, FAM155B may also be able to form a stable complex with NALCN in vivo. This notion is supported by the finding that FAM155A can be functionally substituted by human FAM155B 30.
As our structure was captured as a subcomplex between NALCN and FAM155A, we then asked whether co-expression of NALCN and FAM155A only was also able to recapitulate measurable NALCN current. Interestingly, we have readily recorded large current in response to voltage steps when NALCN and FAM155A was co-transfected (Supplementary Fig. 1c). The current-voltage curve of NALCN co-expressed with either all three auxiliary subunits or with FAM155A only was non-linear (Supplementary Fig. 1d). Compared with NALCN co-expressed with three auxiliary subunits, NALCN with FAM155A exhibited similar voltage sensitivity in current as the apparent gating charge (Zapp) was similar (Supplementary Fig. 1d, Zapp: 1.25 ± 0.55 e0 and 1.22 ± 0.18 e0, respectively). However, the G-V curve of NALCN co-expressed with three auxiliary subunits was largely shifted towards the hyperpolarization voltage as compared to NALCN with FAM155A (V1/2: -44.66 ± 1.91 mV and -65.07 ± 2.11 mV, respectively) (Supplementary Fig. 1e). These findings are distinct from other recent studies 40,41. Moreover, we observed no inactivation of the steady state current by setting the prepulse potential to different levels when NALCN channel was co-expressed with auxiliary subunits (Supplementary Fig. 1f and 1g), which is consistent with the physiological role of NALCN being a leaky sodium channel.
Structural basis for the voltage dependence of NALCN
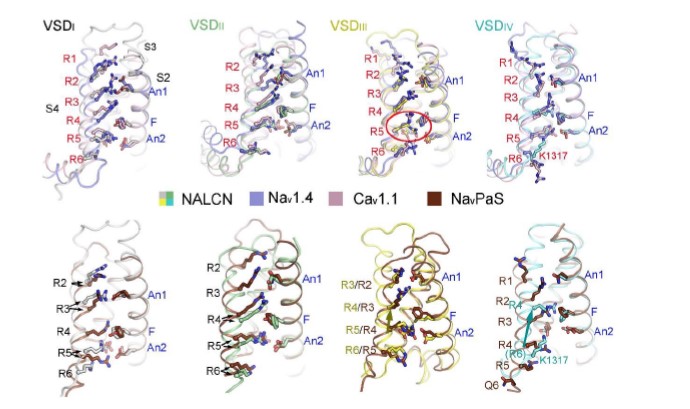
Our structural study has provided an opportunity to understand the voltage sensitivity of NALCN. Detailed analysis of the voltage sensing domains (VSDs) of NALCN suggest that they possess several key features shared among functional VSDs in Nav/Cav channels. For instance, each VSD preserves the gating charges (GCs) in a pattern of occurring every three amino acids in the S4 segments. As usual, we define the position of last gating charge in each S4 as R6 42. NALCN has two to four GCs in each repeat distributed at positions R2-R6 (Fig. 4a). The last GC of repeat IV is at a position that has one residue shift to R6, a phenomenon also observed in other channels 42. Moreover, the residues of charge transfer center (CTC) consists of a negatively charged residue (An2) on S2, an aromatic occluding residue (F) on S2, and a negatively charged residue on S3 are highly conserved in NALCN except for the occluding residue on VSDIII. The An1 sites that are seven residues ahead of the occluding residues on S2 segments of NALCN are mainly negatively charged or polar residues, similar to those of Nav/Cav channels (Fig. 4a, b). In addition, previous structural studies on Nav channels identified several negatively charged or polar residues on S1 that may also play roles in gating charge transfer 34. These residues were also observed on NALCN (Fig. 4a). When the four VSDs are superimposed relative to An1 and CTC, all R4 residues are above the occluding residues on S2, reminiscent of an up or depolarized state of the four VSDs (Fig. 4b, Supplementary Fig. 10).
The voltage sensitivity of NALCN, however, seems to be weaker than that of Nav/Cav channels (Supplementary Fig. 1). Several unique structural features of NALCN may lead to its relative weak voltage sensitivity. First, the occluding residue on S2 in repeat III is replaced by a methionine (Met929), instead of a conserved phenylalanine or tyrosine (Fig. 4c). The occluding residue of the CTC is crucial for the gating transfer during voltage sensing. A previous study shows that mutations of the occluding residue from bulky hydrophobic residue to methionine on a voltage-gated potassium channel diminish its voltage sensitivity 43. Second, the S4 of VSDs are usually formed as 310 helices, whereas in repeat IV the S4 segment largely relax to regular α helix. The only 310 helix turn in S4IV, which is at position of R5, is a serine instead of a GC. These structural observations are also consistent with reported experimental data. For example, it was reported that repeat III and IV contribute little to the voltage sensitivity of NALCN, probably due to their lack of an occluding residue on S2, and lack of GCs and 310 helices on S4, respectively. In contrast, neutralization of the GCs R146+R152 in repeat I or R481+R484 or R481+K487 in repeat II lead to significantly changes in voltage sensitivity 30. Third, the upper gating charges (R2-R4) in repeat I-III form more extensive hydrogen bonding interactions with surrounding negatively charged or polar residues, compared with their counterparts in Nav/Cav channels (Fig. 4c). During the gating charge transfer process, the GCs need to break old interactions with surrounding residues and form new interactions with other residues, accompanying with a sliding movement along the S4 segment. The extensive interactions of the GCs and surrounding residues in NALCN may need the VSDs to overcome a higher energy barrier to undergo conformational changes thus resulting in a weaker voltage sensitivity. Fourth, the S4-5I linker unexpectedly forms a flexible loop instead of a juxtamembrane helix seen in other repeats and other reported Nav/Cav structures. Meanwhile, S5I is extended and elongated to the cytoplasmic region with three helix turns. The unique structure of S4-5I and S5I results in a cavity that accommodate a lipid, that may contribute to the regulation of NALCN (Fig. 2c, Supplementary Fig. 4b). As S4-S5 is supposed to be a key transducer from VSDs to the ion conducing pore during voltage sensing gating in VGICs 44, its relaxation to a flexible linker may hinder the effective transduction of electromechanical coupling of repeat I in NALCN. Altogether, these structural variations in NALCN may contribute to its weak voltage sensitivity.
EEKE selectivity filter in NALCN
Like all reported Nav/Cav structures, the pore domain of NALCN is formed by S5 and S6 helices of the four repeats and the selectivity filter (SF) of NALCN was supported by two pore helices (P1 and P2) that intervene between S5 and S6 (Fig. 5a). The key residues in the SF of NALCN responsible for the ion selectivity are E280/E554/K1115/E1389 (EEKE), distinct from DEKA in Nav channels and EEEE/EEDD in Cav channels. Sequence alignment of the P1-SF-P2 among NALCN, Cav1.1 and Nav1.4 reveals an invariant tryptophan in the first residue of P2 and a highly conserved residue (Thr/Ser/Cys) in the last residue of P1, while other residues are not very conserved (Fig. 5b). The overall structure of P1-SF-P2 in NALCN is more closely related to that of Cav1.1 than to that of Nav1.4, with r.m.s.d. of 0.848 Å over 110 Cα atoms and 1.602 Å over 107 Cα atoms, respectively. Both the pore helices and the SF of NALCN align well with those of Cav1.1, whereas the P2s are not aligned well in superimposition between NALCN and Nav1.4 (Fig. 5c). Despite its structural similarity with Cav channels, NALCN was reported to be mainly selective for monovalent cations and could be blocked by extracellular divalent cations 30. In addition, NALCN is responsible for the background sodium leak conductance in hippocampal neurons, indicating that NALCN is selective for Na+ ion in vivo 5.
To better understand Na+ binding and selectivity in the SF, we performed canonical MD simulation to investigate the interaction of membrane-embedded pore domain in 150 mM NaCl. In three independent trials of 200 ns equilibration, we observe three potential ion binding sites, designated Site1-3, in proximity to SF (Fig. 5d). In addition to the glutamic acid residues in SF, the Na+ binding sites are formed by several negatively changed residues from the pore helices, including E275, D558, D561 and D1390. Among the three binding sites, Site1 is most stably located at the middle of SF, while the other two are transient and presumably cation attractants (Fig. 5d, Supplementary Movie 1). The simulation results were reproducible in three parallel runs. Probability statistics suggest that there is an average of two Na+ ions within 4 Å range of the SF residues through time (Fig. 5e). Notably, Site3 formed by E280/E554/D558 (designated EED site) is spatially close to the Na+ binding site formed by a DEE motif in Nav channels (Fig. 5f) 34. However, the sidechains of the EED motif in NALCN are not as closely spaced as the DEE site in Nav channels and do not form a favorable Na+ binding site. On the other hand, Site1, the most occupied binding site, is spatially close to a putative Ca2+ binding site in Cav1.1 and Cav3.1, suggesting that NALCN and Cav channels share similar ion selectivity mechanisms 35,42 (Fig. 5f). The replacement of glutamic acid (E) or aspartic acid (D) with lysine (K) in repeat III from Cav channels to NALCN has shifted the ion selectivity from Ca2+ to monovalent ions. Like Nav channels, the lysine in NALCN favors Na+ ion and blocks Ca2+ permeation through the selectivity filter. Meanwhile, the EEKE SF may still preserve the ability to bind Ca2+ as the structure of SF in repeat I/II/IV is almost identical to that of Cav channels (Fig. 5c). In this sense, extracellular Ca2+ may compete with Na+ in binding to the SF, explaining why Ca2+ is a blocker of NALCN as shown by previous study 30.
Closed intracellular gate of NALCN
The ion permeation path below the SF in NALCN is enclosed by the S6 tetrahelical bundle, a structural feature that is shared in all reported Nav/Cav structures (Fig. 5a, 6a). However, due to the existence of FAM155A, ions can only enter the permeation path from one side, that is above the pore domain of repeat II (Fig. 6a, left). We used HOLE 45 to calculate the radius along the permeation path and compared it with Nav1.4 and Cav1.1, whose intracellular gates are in open and closed state, respectively. Surprisingly, although NALCN is supposed to conduct a ‘leak’ current, the intracellular gate is closed, even tighter than the closed Cav1.1. The narrowest region, sealed by two layers of hydrophobic residues Val, Ile and Leu, is about 10 Å in length along the permeation path with a radius less than 1 Å (Fig. 6a, right). Notably, the lower gate is fully blocked by the III-IV linker, causing the permeation path to enter the cytosol only from the side. Detailed analysis reveals that the III-IV linker is stabilized by extensive hydrogen bonds or polar interactions with S6I and the II-III linker (Fig. 6b). Most of the involved residues are not conserved among Nav/Cav channels, indicating that the local interactions among S6I, the II-III liner and the III-IV linker are highly specific to NALCN, which implies distinct gating mechanism of NALCN (Supplementary Fig. 7).
It is intriguing that the position of the III-IV linker in NALCN is distinct from those of both Nav1.4 and Cav1.1, in different ways. NALCN does not have an IFM motif, a highly conserved motif in the III-IV linker of all Nav channels that is crucial for fast inactivation through an allosteric inhibition mechanism 44,46. In Nav1.4, the III-IV linker is away from the inner pore, and its helix rotates counterclockwise by about 750 but at a similar vertical height compared to that of NALCN (Fig. 6c, top). The helix in the III-IV linker, however, shows a similar orientation but distinct vertical height in Cav1.1. Compared to Cav1.1, the helix in the III-IV linker of NALCN is about 6 Å up shifted towards the extracellular side, probably due to a shorter loop connecting S6III and the helix in the III-IV linker (Fig. 6c, bottom). The II-III linker, which is part of the elongated S6II in Cav1.1, bends towards the center pore. These structural features in NALCN make S6I, II-III linker and III-IV linker form a close contact that tightly seal the inner gate.
Notably, S5II and S6II are much closer to the center pore in NALCN compared with Nav1.4/Cav1.1. The adjacent S4-5II also displays significant conformational changes in NALCN (Fig. 6c). The dramatic differences in the pore domain of repeat II are closely related to the binding of a lipid molecule (Fig. 2c). The lipid, only observed in NALNC, may play an important role in regulating the gating of NALCN.
Our study reports the high-resolution structure of NALCN in complex with the auxiliary subunit FAM155A. The structure reveals unique features of NALCN that help explain its ion selectivity, voltage sensing and specific interaction with auxiliary subunits. It also provides an important framework for comparative investigations of channel properties of related Nav/Cav channels.
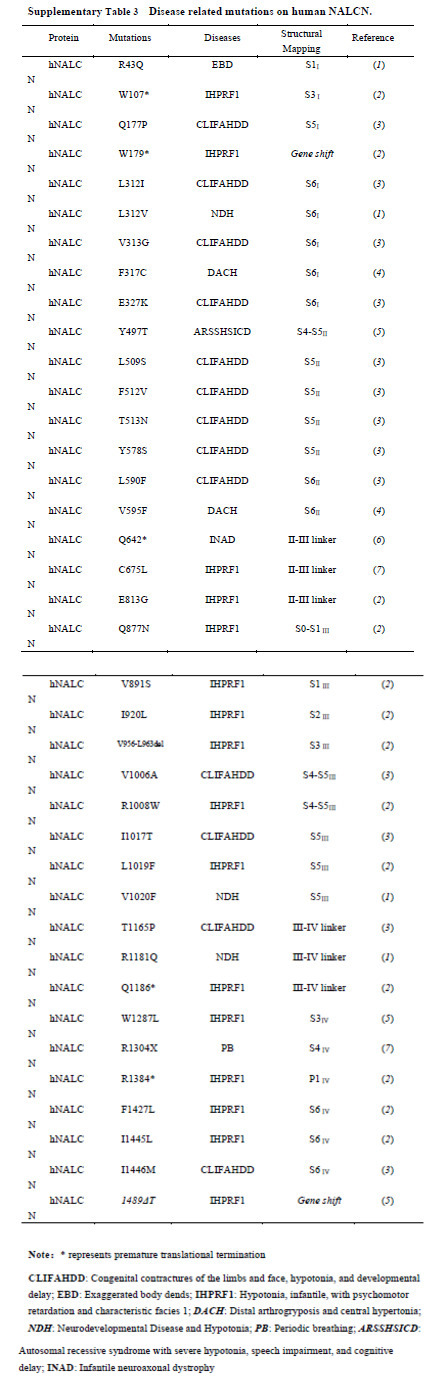
Unlike Nav/Cav channels, there is only one subtype of NALCN in most organisms including human. Certain mutations/deletions of NALCN can be fatal as there are no channels with redundant functions. This idea is supported by the evidence that NALCN is essential for neonatal survival and NALCN knockout mice die within 24 hours after birth 5. We summarized the reported disease-related mutations and mapped them onto the structure (Fig. 7a, Supplementary Table 3). We found that most of the mutations are located on the pore domain of NALCN (Fig. 7a). These mutations may affect NALCN function by directly altering ion permeation properties through the pore. Two mutations on the III-IV linker, T1165P and R1181Q, may lead to changes of channel gating through affecting the local interactions of the III-IV linker with adjacent S6I, the II-III linker, S6IV and CTD. Our structure thus provides an important framework to interpret the potential disease-causing mechanisms of the reported mutations, and shed light on NALCN pharmacology.
TTX is a neurotoxin and sodium channel blocker. The Na+ current conducted by NALCN is TTX-resistant 5. Our structure directly conveys the incompatibility of TTX and the pore of NALCN. Previous structural comparison between the TTX bound Nav1.7 and apo Nav1.5 revealed that a residue with an aromatic ring (Tyr362 in Nav1.7) in repeat I is the structural determinant for TTX sensitivity in Nav channels 38. This residue forms a π-π stacking interaction with TTX bound in the outer pore of Nav1.7 and replacement of the residue with a cystine in TTX-insensitive Nav1.5 results in much lower TTX affinity. The overall backbone of the pore helices and the other residues in contact with TTX are almost the same between Nav1.7 and Nav1.5, though (Fig. 7b). In NALCN, not only the determinant residue is replaced by a glycine, but the other key residues that involved in TTX binding are also not conserved (Supplementary Fig. 8). Moreover, the overall backbone of the pore helices has changed considerably compared with Nav1.7, which lead to clashes within the potential TTX binding site (Fig. 7b). These structural features of NALCN make it unable to bind TTX and even more resistant to TTX than the TTX-insensitive Nav channels.
The relative GCs positions in NALCN VSDs are the same as that of Nav1.4 and Cav1.1, except for the R5 in VSDIII where the side chain of arginine residue faces below the occluding residue in S2. However, the Cα atom of R5 is not shifted, indicating all the VSDs in NALCN are in up conformations (Supplementary Fig. 10). Further comparisons with NavPaS whose VSDIII and VSDIV are in a potential resting state revealed obvious upward movement of the S4 segments in VSDIII and VSDIV from NavPaS to NALCN, confirming the depolarized conformations of VSDs in NALCN (Supplementary Fig. 10). The up VSDs and the closed intracellular gate in NALCN are reminiscent of the inactivated state of Nav/Cav structures 37,42. The significance of such state of NALCN requires further explorations.
The auxiliary subunits play important roles in fine tuning the activity of NALCN. FAM155A, previously characterized as an ER-resident protein in C. elegans 11, turns out to interact with NALCN directly in human. It may help NALCN to fold properly and translocate to cell membrane. Our observation of an evident left shift of the G-V curve upon additional UNC79 and UNC80 co-expressed with NALCN and FAM155A, has also suggested a regulatory effect of UNC79 and UNC80 on NALCN. NALCN contains a π helix in each S6 segment (Fig. 5a). Structural transition between π helix and α helix have been reported in many structures 35,47−49. Association of UNC79, UNC80 into NALCN-FAM155A subcomplex may induce conformational changes and opening of the pore through π-to-α helical transition in S6. Additional components, such as several GPCRs and SFKs may also contribute to the modulation of NALCN function. It remains an open question whether there are extra auxiliary subunits of the NALCN channelosome. Future structural and functional studies on larger complexes of NALCN will deepen our understanding of the regulation of the NALCN channelosome.
While this manuscript was finalizing, Kschonsak et al. reported similar findings in Nature 41 as an Accelerated Article Preview. The back-to-back works mutually verified the main conclusion, and ours also provides several novel points as described.
Protein expression and purification
NALCN, UNC-80, UNC-79 and FAM155A were cloned into pCAG vector. HEK 293F cells (Invitrogen) were transfected with the four plasmids and harvested after 60 hours. Cells were resuspended in buffer containing 25 mM Mops (pH 7.4), 250 mM NaCl, 0.5% (w/v) Lauryl Maltose Neopentyl Glycol (LMNG, Anatrace), 0.06% (w/v) cholesteryl hemisuccinate Tris salt (CHS, Anatrace) and protease inhibitor cocktail including 2 mM phenylmethylsulfonyl fluoride (PMSF), 1.3 µg/ml aprotinin, 0.7 µg/ml pepstatin, and 5 µg/ml leupeptin, then incubated at 4 °C for 2 hours. The insoluble fraction was precipitated by ultracentrifugation at 255,700 g for 1 h, and the supernatant was applied to anti-Flag M2 affinity resin (GenScript) by gravity. The resin was rinsed several times with the Wash buffer, which contains 25 mM Mops (pH 7.4), 500 mM NaCl, 0.01% (w/v) Lauryl Maltose Neopentyl Glycol, and the protease inhibitor cocktail. The target proteins were eluted by the Elution buffer containing 25 mM Mops (pH 7.4), 500 mM NaCl., 0.02% (w/v) glyco diosgenin (GDN, Anatrace), and the protease inhibitor cocktail plus 200 µg/ml FLAG peptide (GenScript). The eluent was then concentrated using a 50-kDa cut-off Centricon (Millipore) and further purified by Superose 6 Increase 10/300 GL column (GE Healthcare) in 25 mM Mops (pH 7.4) 150 mM NaCl, 0.02% (w/v) GDN, and the protease inhibitor cocktail. Fractions were collected and concentrated to about 8.5 mg/ ml for cryo-EM analysis.
Whole-cell electrophysiology
Patch-clamp recordings were performed with a HEKA EPC10 amplifier with PatchMaster software (HEKA) in whole-cell configuration. Patch pipettes were prepared from borosilicate glass and fire-polished to resistance of ~ 6 MΩ. For whole cell recording, serial resistance was compensated by at least 65%. Extracellular solution contained 150 mM NaCl, 10 mM Hepes, and 30 mM d-(+)-glucose (pH 7.4) with NaOH, and intracellular solution contained 136 mM NaCl, 5 mM EGTA, 10 mM Hepes, and 2 mM Na2ATP (adenosine 5′-triphosphate) (pH 7.2) with NaOH. Current amplitude at the steady state during the last 20 ms of voltage steps was averaged to construct the G-V curve. All recordings were performed under room temperature (~22°C). Temperature variation was less than 1 °C as monitored by a thermometer. Current signal was sampled at 20 kHz.
HEK-293T cells were grown in Dulbecco’s modified Eagle’s medium (Thermo Fisher Scientific) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin at 37°C in a 5% CO2 humidified growth incubator. Cells were transiently transfected by Lipofectamine 3000 (Life Technologies) 48-72 hours before patch-clamp recordings. The NALCN, UNC-79, UNC-80, and FAM155A cDNAs were mixed in the mass ratio of 1:1:1:1.
To apply solutions containing ligands such as verapamil during patch-clamp recording, a rapid solution changer with a gravity-driven perfusion system was used (RSC-200, Bio-Logic). Each solution was delivered through a separate tube so that there was no mixing of solutions. Pipette tip with a membrane patch was placed directly in front of the perfusion outlet during recording. Each membrane patch was recorded for only once.
Data from patch-clamp recordings were analyzed in Igor Pro (WaveMatrix). To characterize the steady-state G-V curves, a single-Boltzmann function was used:

where G/Gmax is the normalized conductance, V1/2 is the half-activation voltage, qapp is the apparent gating charge and F is Faraday’s constant, R is gas constant, and T is temperature in Kelvin, which is set to 295 K (22°C).
Cryo-EM sample preparation and data collection
The purified NALCN complex was concentrated to about 8.5 mg/ml for cryo-sample preparation. Aliquots (3.5 μl) of protein solution were loaded onto glow-discharged holey carbon grids (Quantifoil Au R1.2/1.3), which were blotted for 3.5 s and immersed in liquid ethane cooled by liquid nitrogen using Vitrobot (Mark IV, Thermo Fisher Scientific). The grids were exposed through Titan Krios operating at 300 kV equipped with Gatan K3 Summit detector and GIF Quantum energy filter in super-resolution mode (81,000× magnification). Movie stacks were automatically acquired using AutoEMation (Lei et al., 2005), with a 20 eV slit width and a defocus range from -0.5 μm to -2.5 μm. Each stack consisting of 32 frames was exposed for 2.56 s with 0.08 s per frame, and for approximately 50 e-/Å2 of total dose.
Cryo-EM data processing
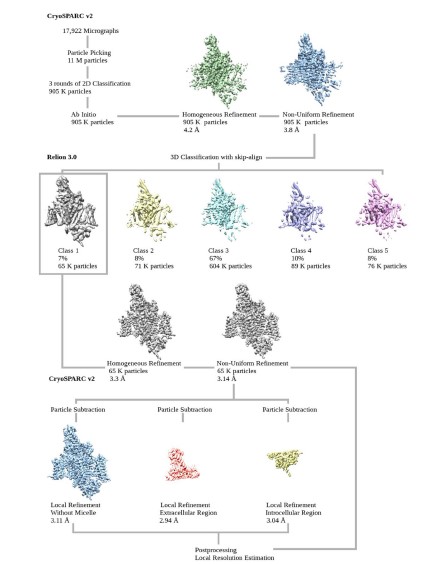
Collected movie stacks were motion-corrected by MotionCor2 50 with 2-fold binning, producing micrographs of 1.087 Å pixel size. Following patch-CTF estimation, around eleven million particles were automatically picked with cryoSPARC v2 51 from 17,922 micrographs. Three rounds of 2D classification enriched images of good classes, resulting in a total of 905,458 particles being selected. An Ab-initio reconstruction generated a low-resolution initial map for the subsequent homogeneous and non-uniform refinement jobs 52, yielding a 3.8-Å reconstruction with identifiable side-chain densities in NALCN pore domain only. To improve map quality, we exported the aligned particles and did a 3D classification without orientation assignment using RELION 3.0 (--skip-align flag) 53,54. One out of five 3D classes represented clear secondary structures of the overall channel complex, and the 65,177 particles were fed back to cryoSPARC v2 for further homogeneous and non-uniform refinements that pushed the resolution to 3.14 Å. Adapted masks for protein regions, extracellular subunit, and intracellular domain were applied in particle subtraction and local refinement tasks, which helped in achieving slightly higher resolution at 3.11 Å, 2.94 Å and 3.04 Å, respectively. Map resolutions were determined by the gold-standard Fourier shell correlation (FSC) 0.143 criterion using Phenix.mtriage 55.
Model building and refinement
An initial model of NALCN was generated using SWISS-MODEL online server 56. The template used for the homology modeling was the α1 subunit of Cav1.1 (PDB: 5GJV). The model was firstly docked into the final reconstruction map in Chimera 57 and then manually adjusted in COOT 58. After building the model of NALCN, an extra density remained unmodelled which is close to the ECL of NALCN in the extracellular side. The high quality of map enables us to identify that the extra density belongs to FAM155A. The structure of FAM155A was then de novo built. Sequence assignment was guided by bulky residues such as Phe, Tyr and Trp. Up to six disulfide bonds and a glycosylation site (Asn217) were identified in FAM155A, further verifying the accuracy of the model. Additional lipid and detergent molecules were manually built to fit into the corresponding densities. In total, 1295 and 182 residues were built for NALCN and FAM155A respectively. The side chains of most of the built residues were assigned except for a short α helix at the C-terminus of NALCN. This short helixis disconnected to any other part of the structure while it is in close contact with the CTD. We tentatively assigned it as part of the CTD (1688-1704) based on sequence and local density analysis. However, there is a possibility that this helix may belong to other interaction proteins such as UNC80, which are not resolved in our current structure.
Subsequently, the models were refined against the corresponding maps by PHENIX 55 in real space (phenix.real_space_refine) with secondary structure and geometry restraints generated by ProSMART 59. Overfitting of the overall models was monitored by refining the models in one of the two independent half maps from the gold-standard refinement approach and testing the refined model against the other map 60. Statistics of 3D reconstruction and model refinement can be found in Supplementary Table 2.
Molecular dynamics simulation
The pore domain of NALCN (residue 171-204, 260-328, 490-603, 1001-1042, 1094-1163, 1318-1400, and 1420-1455) was embedded in 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocholine (POPC) bilayer for molecular dynamics simulations. After determination of side-chain protonation states by PROPKA3.1 61 and peptide terminal neutralization by acetyl/methylamine capping groups, water molecules and 150 mM NaCl were added using VMD 62, resulting in a system of size 100 × 100 × 105 Å with ~93,000 atoms. The system was parameterized using tleap in AmberTools1863 by ff14SB 64 and LIPID17 force fields, and along with TIP3P water model65. All the simulations were performed using OpenMM7 66. We first applied positional constraints (k=10 kcal/mol/Å2) on protein heavy atoms for a 5000-step minimization, then released constraints of side-chains when heating up the system to 310 K with 2 fs stepsize (H-bonds constraints), 1 ps-1 friction coefficient for Langevin dynamics, Particle Mesh Ewald (PME) method 67, and 12 Å non-bonded cutoff. During 50-ns pre-equilibration for membrane relaxation in NPT ensemble, the force constant k of backbone atoms was reduced from 10 to 0.1 kcal/mol/Å2. Three independent 200-ns production runs were conducted constraining Cα atoms more than 15 Å away from the selectivity filter residues (EEKE) with k = 0.1 kcal/mol/Å2. Simulation trajectories were analyzed using MDTraj 1.9.3 68. After superimposition of the pore region to the initial structure, we recorded the positions of sodium ions within 15 Å of SF residues EEKE every 1 ns for each run.
Protein crosslinking and LC-MS/MS analysis
Equal amount (w/w) of BS3 (bis[sulfosuccinimidyl] suberate) was added into the protein mixture, which was incubated at room temperature for 1 h. The reaction was terminated by adding 0.5 M ammonium bicarbonate to a final concentration of 20 mM for 10 min incubation. The SDS-PAGE was used to separate the crosslinked protein and stained with Coomassie Blue G-250. The gel bands of interest were cut into pieces. Sample was digested by trypsin with prior reduction and alkylation in 50 mM ammonium bicarbonate at 37ºC overnight. The digested products were extracted twice with 1% formic acid in 50% acetonitrile aqueous solution and dried to reduce volume by speedvac.
For LC-MS/MS analysis, the peptides were separated by a 65 min gradient elution at a flow rate 0.300 µL/min with the Thermo EASY-nLC1200 integrated nano-HPLC system which is directly interfaced with the Thermo Q Exactive HF-X mass spectrometer. The analytical column was a home-made fused silica capillary column (75 µm ID, 150 mm length; Upchurch, Oak Harbor, WA) packed with C-18 resin (300 A, 3 µm, Varian, Lexington, MA). Mobile phase A consisted of 0.1% formic acid, and mobile phase B consisted of 100% acetonitrile and 0.1% formic acid. The mass spectrometer was operated in the data-dependent acquisition mode using the Xcalibur 4.1 software and there is a single full-scan mass spectrum in the Orbitrap (300-1800 m/z, 60,000 resolution) followed by 20 data-dependent MS/MS scans at 30% normalized collision energy. Each mass spectrum was analyzed using the Thermo Xcalibur Qual Browser and pLink 2 69 for the database searching and crosslinking analysis.
Author Contributions.
Z.Y. conceived the project. Z.Y., J.W. designed all experiments and supervised the project. J.X., and Z.Y. prepared the protein sample, J.W., M.K. and J.X. collected cryo-EM data and calculated the cryo-EM map. J.W. built the atomic model. M.K. performed the MD simulations. F.Y. designed and L.X. conducted the electrophysiological experiments. All authors contributed to data analysis. Z.Y. and J.W. wrote the manuscript.
Acknowledgements.
We thank the Cryo-EM Facility of Westlake University for providing cryo-EM and computation support. We thank Westlake University Supercomputer Center for computational resource and related assistance. We thank the Mass Spectrometry & Metabolomics Core Facility at the Center for Biomedical Research Core Facilities of Westlake University for MS sample analysis. We thank Dr. Stephan Pless (University of Copenhagen) for kindly sharing the plasmids of human UNC-79, UNC-80 and FAM155A. We thank Dr. Hongtao Yu (Westlake University), Dr. Nieng Yan (Princeton University) for critical reading of the manuscript. This work was supported by Westlake Education Foundation. Atomic coordinate and corresponding EM maps of the NALCN-FAM155A complex (PDB: 7CM3; EMDB: EMD-30400) have been deposited in the Protein Data Bank (http://www.rcsb.org) and the Electron Microscopy Data Bank (https://www.ebi.ac.uk/pdbe/emdb/), respectively. Correspondence should be addressed to Z. Yan ([email protected]), J. Wu ([email protected]), or F. Yang ([email protected]).
- Bean, B.P. The action potential in mammalian central neurons. Nature Reviews Neuroscience 8, 451–465 (2007).
- Ren, D.J. Sodium Leak Channels in Neuronal Excitability and Rhythmic Behaviors. Neuron 72, 899–911 (2011).
- Catterall, W.A. The molecular basis of neuronal excitability. Science 223, 653 – 61 (1984).
- Lee, J.H., Cribbs, L.L. & Perez-Reyes, E. Cloning of a novel four repeat protein related to voltage-gated sodium and calcium channels. FEBS Lett 445, 231–6 (1999).
- Lu, B. et al. The neuronal channel NALCN contributes resting sodium permeability and is required for normal respiratory rhythm. Cell 129, 371–83 (2007).
- Jospin, M. et al. UNC-80 and the NCA ion channels contribute to endocytosis defects in synaptojanin mutants. Curr Biol 17, 1595–600 (2007).
- Lear, B.C. et al. UNC79 and UNC80, putative auxiliary subunits of the NARROW ABDOMEN ion channel, are indispensable for robust circadian locomotor rhythms in Drosophila. PLoS One 8, e78147 (2013).
- Yeh, E. et al. A putative cation channel, NCA-1, and a novel protein, UNC-80, transmit neuronal activity in C. elegans. PLoS Biol 6, e55 (2008).
- Lu, B. et al. Extracellular calcium controls background current and neuronal excitability via an UNC79-UNC80-NALCN cation channel complex. Neuron 68, 488 – 99 (2010).
- Wang, H. & Ren, D. UNC80 functions as a scaffold for Src kinases in NALCN channel function. Channels 3, 161–163 (2009).
- Xie, L. et al. NLF-1 Delivers a Sodium Leak Channel to Regulate Neuronal Excitability and Modulate Rhythmic Locomotion. Neuron 77, 1069–1082 (2013).
- Swayne, L.A. et al. The NALCN ion channel is activated by M3 muscarinic receptors in a pancreatic beta-cell line. Embo Reports 10, 873–880 (2009).
- Lu, B.X. et al. Peptide neurotransmitters activate a cation channel complex of NALCN and UNC-80. Nature 457, 741-U10 (2009).
- Senatore, A., Monteil, A., van Minnen, J., Smit, A.B. & Spafford, J.D. NALCN Ion Channels Have Alternative Selectivity Filters Resembling Calcium Channels or Sodium Channels. Plos One 8(2013).
- Eigenbrod, O. et al. Rapid molecular evolution of pain insensitivity in multiple African rodents. Science 364, 852–859 (2019).
- Nash, H.A., Scott, R.L., Lear, B.C. & Allada, R. An unusual cation channel mediates photic control of locomotion in Drosophila. Current Biology 12, 2152–2158 (2002).
- Lear, B.C. et al. The ion channel narrow abdomen is critical for neural output of the Drosophila circadian pacemaker. Neuron 48, 965–976 (2005).
- Chong, J.X. et al. De novo mutations in NALCN cause a syndrome characterized by congenital contractures of the limbs and face, hypotonia, and developmental delay. Am J Hum Genet 96, 462–73 (2015).
- Fukai, R. et al. De novo missense mutations in NALCN cause developmental and intellectual impairment with hypotonia. J Hum Genet 61, 451–5 (2016).
- Bend, E.G. et al. NALCN channelopathies: Distinguishing gain-of-function and loss-of-function mutations. Neurology 87, 1131–1139 (2016).
- Al-Sayed, M.D. et al. Mutations in NALCN cause an autosomal-recessive syndrome with severe hypotonia, speech impairment, and cognitive delay. Am J Hum Genet 93, 721–6 (2013).
- Angius, A. et al. Novel NALCN biallelic truncating mutations in siblings with IHPRF1 syndrome. Clin Genet 93, 1245–1247 (2018).
- Bramswig, N.C. et al. Genetic variants in components of the NALCN-UNC80-UNC79 ion channel complex cause a broad clinical phenotype (NALCN channelopathies). Human Genetics 137, 753–768 (2018).
- Koroglu, C., Seven, M. & Tolun, A. Recessive truncating NALCN mutation in infantile neuroaxonal dystrophy with facial dysmorphism. J Med Genet 50, 515–20 (2013).
- Askland, K., Read, C. & Moore, J. Pathways-based analyses of whole-genome association study data in bipolar disorder reveal genes mediating ion channel activity and synaptic neurotransmission. Hum Genet 125, 63–79 (2009).
- Mok, K.Y. et al. Genomewide association study in cervical dystonia demonstrates possible association with sodium leak channel. Mov Disord 29, 245–51 (2014).
- Wang, K.S., Liu, X.F. & Aragam, N. A genome-wide meta-analysis identifies novel loci associated with schizophrenia and bipolar disorder. Schizophr Res 124, 192–9 (2010).
- Shamseldin, H.E. et al. Mutations in UNC80, Encoding Part of the UNC79-UNC80-NALCN Channel Complex, Cause Autosomal-Recessive Severe Infantile Encephalopathy. Am J Hum Genet 98, 210–5 (2016).
- Stray-Pedersen, A. et al. Biallelic Mutations in UNC80 Cause Persistent Hypotonia, Encephalopathy, Growth Retardation, and Severe Intellectual Disability. Am J Hum Genet 98, 202–9 (2016).
- Chua, H.C., Wulf, M., Weidling, C., Rasmussen, L.P. & Pless, S.A. The NALCN channel complex is voltage sensitive and directly modulated by extracellular calcium. Science Advances 6(2020).
- Bouasse, M., Impheng, H., Servant, Z., Lory, P. & Monteil, A. Functional expression of CLIFAHDD and IHPRF pathogenic variants of the NALCN channel in neuronal cells reveals both gain- and loss-of-function properties. Scientific Reports 9(2019).
- Catterall, W.A. From ionic currents to molecular mechanisms: the structure and function of voltage-gated sodium channels. Neuron 26, 13–25 (2000).
- Folander, K. et al. Cloning and expression of the delayed-rectifier IsK channel from neonatal rat heart and diethylstilbestrol-primed rat uterus. Proc Natl Acad Sci U S A 87, 2975–9 (1990).
- Pan, X. et al. Structure of the human voltage-gated sodium channel Nav1.4 in complex with beta1. Science 362(2018).
- Zhao, Y.Y. et al. Molecular Basis for Ligand Modulation of a Mammalian Voltage-Gated Ca2 + Channel. Cell 177, 1495-+ (2019).
- Shen, H. et al. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 362(2018).
- Shen, H., Liu, D., Wu, K., Lei, J. & Yan, N. Structures of human Nav1.7 channel in complex with auxiliary subunits and animal toxins. Science 363, 1303–1308 (2019).
- Jiang, D. et al. Structure of the Cardiac Sodium Channel. Cell 180, 122–134 e10 (2020).
- Holm, L. DALI and the persistence of protein shape. Protein Science (2019).
- Chua, H.C., Wulf, M., Weidling, C., Rasmussen, L.P. & Pless, S.A. The NALCN channel complex is voltage sensitive and directly modulated by extracellular calcium. Sci Adv 6, eaaz3154 (2020).
- Kschonsak, M. et al. Structure of the human sodium leak channel NALCN. Nature (2020).
- Wu, J.P. et al. Structure of the voltage-gated calcium channel Ca(v)1.1 at 3.6 angstrom resolution. Nature 537, 191-+ (2016).
- Tao, X., Lee, A., Limapichat, W., Dougherty, D.A. & MacKinnon, R. A gating charge transfer center in voltage sensors. Science 328, 67–73 (2010).
- Yan, Z. et al. Structure of the Nav1.4-beta1 Complex from Electric Eel. Cell 170, 470–482 e11 (2017).
- Smart, O.S., Neduvelil, J.G., Wang, X., Wallace, B.A. & Sansom, M.S.P. HOLE: A program for the analysis of the pore dimensions of ion channel structural models. Journal of Molecular Graphics & Modelling 14, 354-& (1996).
- Armstrong, C.M. & Bezanilla, F. Inactivation of the sodium channel. II. Gating current experiments. J Gen Physiol 70, 567–90 (1977).
- McGoldrick, L.L. et al. Opening of the human epithelial calcium channel TRPV6. Nature 553, 233-+ (2018).
- Singh, A.K., Saotome, K., McGoldrick, L.L. & Sobolevsky, A.I. Structural bases of TRP channel TRPV6 allosteric modulation by 2-APB. Nature Communications 9(2018).
- Su, Q. et al. Cryo-EM structure of the polycystic kidney disease-like channel PKD2L1. Nature Communications 9(2018).
- Zheng, S.Q. et al. MotionCor2: anisotropic correction of beam-induced motion for improved cryo-electron microscopy. Nature Methods 14, 331–332 (2017).
- Punjani, A., Rubinstein, J.L., Fleet, D.J. & Brubaker, M.A. cryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nature Methods 14, 290-+ (2017).
- Ali Punjani, H.Z., David J. Fleet. Non-uniform refinement: Adaptive regularization improves single particle cryo-EM reconstruction. bioRxiv (2019).
- Zivanov, J. et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. Elife 7(2018).
- Scheres, S.H.W. RELION: Implementation of a Bayesian approach to cryo-EM structure determination. Journal of Structural Biology 180, 519–530 (2012).
- Adams, P.D. et al. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallographica Section D-Structural Biology 66, 213–221 (2010).
- Waterhouse, A. et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Research 46, W296-W303 (2018).
- Pettersen, E.F. et al. UCSF chimera - A visualization system for exploratory research and analysis. Journal Of Computational Chemistry 25, 1605–1612 (2004).
- Emsley, P. & Cowtan, K. Coot: model-building tools for molecular graphics. Acta Crystallographica Section D-Biological Crystallography 60, 2126–2132 (2004).
- Nicholls, R.A., Fischer, M., McNicholas, S. & Murshudov, G.N. Conformation-independent structural comparison of macromolecules with ProSMART. Acta Crystallogr D Biol Crystallogr 70, 2487–99 (2014).
- Amunts, A. et al. Structure of the yeast mitochondrial large ribosomal subunit. Science 343, 1485–1489 (2014).
- Olsson, M.H., Sondergaard, C.R., Rostkowski, M. & Jensen, J.H. PROPKA3: Consistent Treatment of Internal and Surface Residues in Empirical pKa Predictions. J Chem Theory Comput 7, 525–37 (2011).
- Humphrey, W., Dalke, A. & Schulten, K. VMD: visual molecular dynamics. J Mol Graph 14, 33 – 8, 27 – 8 (1996).
- D.A. Case et al. AMBER 2018. (University of California, San Francisco, 2018).
- Maier, J.A. et al. ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from ff99SB. J Chem Theory Comput 11, 3696–713 (2015).
- Jorgensen, W.L., Chandrasekhar, J., Madura, J.D., Impey, R.W. & Klein, M.L. Comparison of simple potential functions for simulating liquid water. The Journal of Chemical Physics 79, 926–935 (1983).
- Eastman, P. et al. OpenMM 7: Rapid development of high performance algorithms for molecular dynamics. Plos Computational Biology 13(2017).
- Essmann, U. et al. A smooth particle mesh Ewald method. The Journal of Chemical Physics 103, 8577–8593 (1995).
- McGibbon, R.T. et al. MDTraj: A Modern Open Library for the Analysis of Molecular Dynamics Trajectories. Biophys J 109, 1528–32 (2015).
- Chen, Z.L. et al. A high-speed search engine pLink 2 with systematic evaluation for proteome-scale identification of cross-linked peptides. Nat Commun 10, 3404 (2019).
- Goddard, T.D. et al. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Science 27, 14–25 (2018).
- DeLano, W.L. The PyMOL Molecular Graphics System on World Wide Web http://www.pymol.org.. (2002).
There is NO Competing Interest.
- movie.mp4
Supplementary Movie 1 MD simulation of NALCN pore domain for sodium binding site identification. Left panel. Top view of selectivity filter. Ion binding residues are highlighted in stick representation. Probability density of sodium is illustrated in mesh grid. Sodium ions are colored in magenta. Right panel. Side views of selectivity filter.
- NALCNSI.docx
Supplementary information
- ZhenFLATR.pdf
Reporting Summary
- ZhenPDB.pdf
PBD validation report
- FigureS1.jpg
. Electrophysiological properties of NALCN characterized by patch-clamp recordings. (a) Representative whole-cell recordings of HEK293 with mock transfection and NALCN channel transfected, respectively. Dashed line indicates the zero-current level. (b) Representative whole-cell recordings of NALCN, FAM155A, UNC79 and UNC80 co-expressed in HEK293 cells. Data points in shaded rectangle were averaged for plotting current-voltage curve in (d). Dashed line in black indicates the zero-current level. Dashed curves in red and blue are fitted curves to the inactivation and activation kinetics with exponential functions, respectively. NALCN current was inhibited by 1 mM verapamil and the current could be recovered after washing off. The same whole-cell patch was used during verapamil inhibition. (c) Representative whole-cell recordings of NALCN channel and FAM155A co-expressed in HEK293 cells. Data points in shaded rectangle were averaged for plotting current-voltage curve. (d) Current-voltage curve of NALCN co-expressed with FAM155A only (n = 6) and co-expressed with FAM155A, UNC79 and UNC80 (n = 7). The currents were normalized to the maximum current level. (e) Conductance-voltage (G-V) curves of NALCN co-expressed with FAM155A (Red: V1/2 = -44.66 ± 1.91 mV; apparent gating charge (Zapp) = 1.22 ± 0.18 e0 ) and co-expressed with FAM155A, UNC79 and UNC80 (Blue: V1/2 = -65.07 ± 2.11 mV; apparent gating charge (Zapp) = 1.25 ± 0.55 e0 ). G-V curves were fitted to a single-Boltzmann function (equation (1) in Methods, dash curves). Co-expression with FAM155A, UNC79 and UNC80 shifted the G-V curve of NALCN channel towards the hyperpolarization voltage. All data are given as mean ± s.e.m. (f) Representative whole-cell recordings of the steady-state inactivation of NALCN co-expressed with FAM155A (left) or with FAM155A, UNC79 and UNC80 (right). (g) Prepulse voltage dependence of steady-state inactivation of NALCN channel co-expressed with auxiliary subunits measured from -80 mV (n = 7).
- FigureS2.jpg
Cryo-EM analysis of human NALCN in complex with FAM155A. (a) Last step purification of recombinantly expressed NALCN-FAM155A complex. A representative chromatogram of gel filtration purification is shown. The indicated fractions were resolved by SDS-polyacrylamide gel electrophoresis followed by Coomassie blue staining. The bands corresponding to NALCN and FAM155A are indicated by black and orange arrows, respectively. The protein identity of each band was confirmed by mass spectrometric analysis. (b) Representative micrograph of the NALCN-FAM155A complex. Scale bar, 40 nm. (c) Representative two-dimensional class averages for NALCN-FAM155A complex. Box size: 240 Å; circle mask: 220 Å. (d) Angular distribution of the particles of the final reconstruction generated by cryoSPARC. (e) Gold standard FSC curves for the 3D reconstructions. The curves for the reconstructions of the overall map, map of the intracellular region and map of the extracellular region are indicated by blue, yellow, and red lines, respectively. (f) Validation of the final structure models. FSC curves of the final refined model versus the corresponding map that it was refined against (black); of the model refined in the first of the two independent half maps used for the gold-standard Fourier shell correlation curves versus that same map (blue); and of the model refined in the first of the two independent maps versus the second independent map (red). The small difference between the blue and red curves indicates that the refinement of the atomic coordinates did not suffer from overfitting.
- FigureS3.jpg
Flowchart for EM data processing. Please refer to “Image processing” session in Methods for details.
- FigureS4.jpg
EM maps of human NALCN. (a) EM maps for the S1-S6 segments in the four repeats of NALCN. The side groups of representative bulky residues are labeled. The gating charges in S4 helices are highlighted in brown. The densities, shown as blue meshes, are contoured at 4-5 σ in PyMol. (b) EM maps for the two resolved lipid molecules that located in the inner side of the membrane. Two perpendicular views are shown for each lipid. The lipid densities are contoured at 3-4 σ in PyMol.
- FigureS5.jpg
EM maps of NALCN-FAM155A complex. (a) All residues that constitute the SF in four repeats of NALCN are clearly resolved. The densities are contoured at 4-5 σ in PyMol. The EEKE residues are labeled orange. (b) Representative EM maps of FAM155A. The side groups of bulky residues are labeled. (c) EM maps of the disulfide bonds in the complex structure. The densities are contoured at 3-4 σ in PyMol. (d) EM maps of the sugar moieties in the glycosylation sites. The densities are contoured at 2-4 σ in PyMol.
- FigureS6.jpg
Mass Spectrometric analysis of crosslinked NALCN-FAM155A complex. Shown here is a high resolution HCD spectra of the inter-subunit crosslinked peptides from NALCN-FAM155A complex. The number shown in the brackets refer to the residue number in the indicated subunit. Inset: Structure of the NALCN-FAM155A interaction interface. K1377 of NALCN is shown in stick. The C-terminal residue in the resolved FAM155A structure is E382, which is about 27 Å from K1377 of NALCN. K395 of FAM155A, 13 residues after E382, is likely to be in the vicinity of cross-linking range with K1377 of NALCN. The cross-linking MS result is consistent with our structure observation.
- FigureS7.jpg
Interaction interfaces between NALCN and FAM155A. (a) Structure of the FAM155A. The N-lobe and C-lobe were colored in gold and orange, respectively. The secondary elements are labeled based on the definition in Figure 3. (b) Crystal structure of the CRD domain of MuSK and its superimposition with FAM155A. The conserved disulfide bonds between the two structures are highlighted by black arrows. (c) Electrostatic surface of the interaction interfaces between NALCN and FAM155A. The structures are shown with both subunits in electrostatic surface (left), with NALCN in cartoon and FAM155A in electrostatic surface (middle), or with FAM155A in cartoon and NALCN in electrostatic surface (right). Two side views are presented. The green circles highlight the interaction interfaces that have complementary electrical potential between NALCN and FAM155A.
- FigureS8final2.jpg
Sequence alignment among NALCN and selected Nav/Cav channels. The primary sequences of four human Nav subtypes and three human Cav subtypes are compared with that of human NALCN using Clustal W (8). The invariant residues are shaded gray and the conserved residues are colored red. Secondary structural elements of human NALCN are presented above the sequence alignment and color-coded for each repeat. The critical EEKE residues in the selectivity filter are colored red and shaded yellow. The gating charge residues (labeled R1-R6) in each S4 segments are colored white and shaded red. The residues on S1-S3 that may facilitate the gating charge transfer, including the labeled An1 and An2 sites and the occluding residues on S2, are shaded cyan. The residues that form disulfide bonds are labeled with squares below the sequence alignment. The glycosylation sites are labeled with black triangles and the residues that mediate interaction with FAM155A are labeled with orange triangles. The Uniprot IDs for the aligned sequences are: hNALCN: Q8IZF0; hNav1.2:Q99250; hNav1.4:P35499; hNav1.5:Q14524; hNav1.7:Q15858; hCav1.1: Q13698; hCav2.1: O00555; hCav3.1: O43497.
- FigureS9.jpg
Sequence alignment of FAM155A. The primary sequences of FAM155A from human, mouse, and bovine and FAM155B from human are aligned using Clustal W. The invariant residues are colored white and shaded gray. The conserved residues are colored orange. The secondary structure elements of human FAM155A are indicated above the sequence alignment. The residues that form disulfide bonds are labeled with squares below the sequence alignment. The glycosylation site is labeled with black triangle and the residues that mediate interaction with NALCN are labeled with orange triangles. The Uniprot IDs for the aligned sequences are: hFAM155A: B1AL88, mFAM155A: Q8CCS2; bFAM155A: A4IFM1, hFMA155B:O75949.
- FigureS10.jpg
All four VSDs adopt up conformations. Each VSD of NALCN is superimposed with their counterpart in Nav1.4 (PDB: 6AGF) and Cav1.1 (6JP5). The VSDs are aligned relative to CTC residues and An1 on S2 (labeled blue). S1 segments were omitted by visual clarity. The gating charges (GCs), charge transfer center (CTC) and An1 on S2 are shown as stick and Cα atoms of the GCs are shown as spheres. The GCs on S4 are labeled as R1-R6. The positions of the GCs in NALCN are similar as that of Nav1.4 and Cav1.1, which represent up or depolarized conformations. By contrast, the positions of GCs in VSDIII and VSDIV show an upward shift compared to that of NavPaS, whose VSDIII/IV may represent resting state.
- TableS1.png
Data collection and model statistics
- TableS2.png
Summary of contact interfaces between NALCN and FAM155A
- TableS3final.jpg
Disease related mutations on human NALCN.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}