Donor GFP-labeled stem cells create chimeric spiral ganglion and vestibular neurons in the Neurog1+/- heterozygote
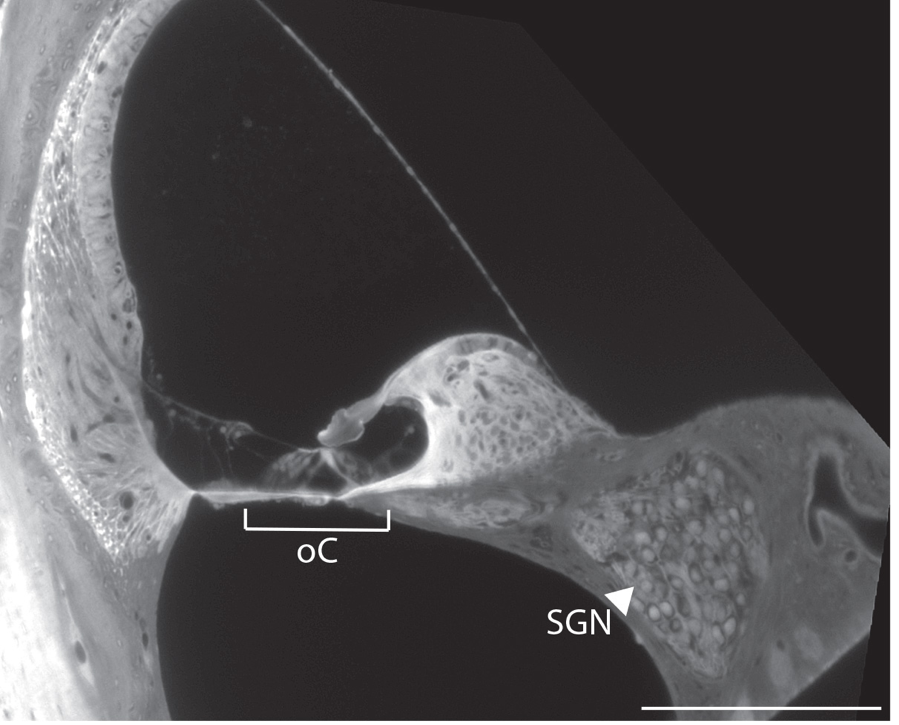
Neurog1-deficient blastocysts were established by performing in vitro fertilization (IFV) with zygotes extracted from Neurog1+/- heterozygous dams and fresh sperm from Neurog1+/- heterozygous male mice. Blastocysts were injected with GFP-labeled mouse induced pluripotent stem cells (iPSCs) at approximately embryonic day (E)3.5 and subsequently were transferred into surrogate pseudopregnant dams (Fig. 1A). Upon analysis at post-natal day (P)1, robust and specific stem cell incorporation of GFP-labeled cells derived from the donor stem cells was seen in the SGN of a Neurog1+/- heterozygote (Fig. 1).
In the complemented Neurog1+/-, GFP-labeled iPSCs contributed to the SGN and descending neuronal processes in the cochlea with minimal donor stem cell-derived cell incorporation in other tissue (Fig. 1D,E arrowheads). The specificity of labeling in the Neurog1+/- was unmistakable, given the lack of GFP labeling observed in the wild type SGN (Fig. 1C,F, arrows). GFP-labeled donor iPSCs also contributed to the cell bodies of Scarpa’s ganglion (Fig. 1G), in addition to vestibular neurons innervating the cristae ampullaris (Fig. 1H,I). Since Neurog1 is known to be required for the formation of both cochlear and vestibular neurons (16), these results indicate that the integration pattern of cells derived from the wild type donor cells recapitulates that expected from cells wild type endogenous gene function and that Neurog1 haplodeficiency creates a vacent niche that can be filled by cells derived from exogenous stem cells to produce SGNs using BC.
Complementation of Neurog1-deficiency is distinct from general chimerism
A chimera is defined as a composite animal comprised of two genetically distinct cell populations (29). Performing BC in wild type animals will result in random chimerization throughout the developing embryo (30, 31). When performing BC in knockout blastocysts, in addition to exogenous donor-derived cells target a vacant developmental niche, random chimerism can also be observed in non-targeted cell types.
Two examples of complemented Neurog1+/- heterozygotes were highly chimeric, as a high degree of specific donor derived cell integration into the SGN was observed (Fig. 2B-D’), in addition to general chimerism, which was evident from the widespread GFP expression in non-sensory otic cell types (Fig. 2B-D’). This general chimerism is similar to that observed in the Neurog1+/+ control which also displayed the incorporation of GFP-expressing cells in non-sensory cells. Notably, the Neurog1+/- heterozygote SGNs, in some regions, appeared to be derived nearly entirely from donor iPSCs, whereas the Neurog1+/+ wild type SGNs were consistently negative for GFP expression (Figs. 1C, 2A,A’).
The extent of donor-derived cell chimerism in the Neurog1+/+ wild type in comparison to the complemented Neurog1+/- heterozygotes was assessed further by looking at the resepective level of GFP expression in the temporal lobe of the brain. This analysis clearly showed the incorporation of exogenous iPSC-derived cells in the Neurog1+/+ brain at a comparable level to the complemented Neurog1+/- heterozygotes (Fig. 2E,F). Therefore, chimeras successfully formed regardless of geneotype, but in the absence of a Neurog1-deficient niche, no GFP-expressing cells derived from the exogenous donor iPSCs contributed to the formation of the SGN in wild type inner ears (Figs. 1F, 2A’). These results support our hypothesis by demonstrating that BC can specifically generate SGNs using donor stem cells.
We did not see complementation in the two Neurog1-/- mutant embryos that we obtained. However, both nulls showed few to no GFP-expressing cells engrafted throughout the whole embryo, suggesting that the lack of complementation was due to low chimerism. With an increased sample size, we expect to obtain highly chimeric Neurog1-/- mutants in which all of the SGNs are derived from donor stem cell progeny. Therefore, it is anticipated that, following Medel’s law, 75% of blastocysts (heterozygous and null) obtained from BC will give rise to chimeric inner ears.
Donor-derived stem cells extensively contibute to the complemented Neurog1+/- vestibule
In the highly chimeric complemented Neurog1+/- inner ears, the contribution of donor-derived GFP-labeled cells appeared to increase in sections through the more basal cochlea in a trend that was dramatically more evident in the vestibule. In fact, it appeared that the majority of cells in the vestibule were derived from donor iPSCs, given the extensive presence of GFP in all vestibular cell types. Specifically, GFP entirely co-expressed with the neuronal marker class III beta-tubulin (TUJ1) in the neurites innervating the vestibular sensory organs (Fig 3A-B’, arrow head and dotted lines), in addition to many Myosin 6 (MYO6)-expressing vestibular hair cells (Fig 3A-B, red) and nonsensory cells (Fig 3A’-B’, small arrows). While the degree of donor cell contribution to the complemented Neurog1+/- vestibule was striking, the biological significance of this was lacking, until a clear heterozygote effect in non-complemented Neurog1+/- inner ears was detected.
Blastocyst complementation rescues Neurog1+/- inner ear malformations
Non-complemented Neurog1+/- heterozygote inner ears were observed to have inner ear morphological non-sensory malformations, which included inner ears reduced in size by approximately 60% of the wildtype control (Fig 4A, Supplemental Movie 1). Three-dimensional reconstructions revealed overt malformations particularly in the vestibule, in which the anterior and lateral ampullae and the saccule were notably reduced in size (Fig. 4A-D, Supplemental Movie 1). This finding was confirmed via cryosections, which displayed that the non-complemented Neurog1+/- vestibule sometimes had an abnormally orientated saccule, utricle, and anterior ampullae. Specifically, unlike the typical orthogonal arrangement of the utricular and saccular maculae observed in the wild type vestibule, the non-complemented Neurog1+/- sensory maculae (denoted by MYO6-expressing hair cells) were oriented in parallel (Fig. 4E,F; red, small arrows). Additionally, the utricular maculae and anterior crista ampullaris were unusually close to one another (Fig. 4F, arrowheads). Moreover, in some cases, the lateral ampulla appeared connected to the lateral semi circular canal (Fig. 4I, arrowheads). Together these observations suggest that a reduction in non-sensory cell formation and/or a failure of sensory organ separation occured with the reduction of Neurog1 gene dose.
Strikingly, two Neurog1+/- heterozygotes completely lacked a vestibule (not shown). Cochlear and vestibular HCs developed normally in the Neurog1+/- heterozygote (as has been reported for the Neurog1-/- mutant) despite impaired non-sensory formation. However, sensory development potentially occurred at the expense of non-sensory development, as ectopic HCs were sometimes seen in non-sensory regions in the lateral semi circular canal (Fig. 4I’).
Importantly, non-sensory defects observed in the Neurog1+/- heterozygotes were rescued in complemented Neurog1+/- samples (Fig. 4A’-D,G,J). Given the extensive contribution of GFP to nonsensory tissue in the complemented Neurog1+/- heterozygous vestibular sensory organs (Fig. 3), these results suggest that widespread incorporation of cells derived from the donor iPSCs to the vestibule in the complemented Neurog1+/- heterozygote is not random, but rather reflects the recovery of a previously unappreciated biological function of Neurog1 in inner ear morphogenesis. This finding demonstrates the use of BC as a tool to elucidate novel gene function and to confirm or disprove concepts regarding the development of neurobiological systems.
{kind=link}