SARS-CoV-2 NanoBiT biosensor for detecting ACE2-RBD interactions
Several different reporter fragment complementation-based strategies have been employed to interrogate protein-protein interactions13, including split-luciferase schemes14-16 17. Conventional split-luciferase bioreporters can be limited in their application due to their relatively large sizes, poor stability, and the short half-lives of their catalyzed luminescent reactions. The recently reported Nanoluciferase (or NanoLuc from Oplophorus gracilirostris)18 does not possess these limitations, and a NanoLuc-based fragment complementation system has been reported 10, 11 12. Our biosensor employs NanoLuc fragments linked to RBD and ACE2 creating a biosensor that can rapidly and sensitively serve as a surrogate for virus:host cell interactions (Fig. 1A). Using published sequences and structural homology analysis7, 8, 19, 20, we designed a SARS-CoV-2 RBD sequence spanning residues 331 to 524 of the S protein (194 amino acids; Supplementary Fig. 1) for one component of the biosensor. For the other component we used the soluble ectodomain of ACE2 (residues 1 to 740) as this has been shown to be sufficient to interact with RBD21. Since RBD is the smaller protein of the two partners of interest11, we linked the RBD with the larger fragment of the split luciferase (LgBiT) while ACE2 was fused to the small fragment (SmBiT, Figure 1B). A glycine-serine linker was inserted between ACE2 or RBD and its respective NanoBiT component11 to enhance folding and flexibility of the fusion proteins. To facilitate the production of the interacting partners as secreted molecules, we carried out a series of codon optimization studies and tested different secretion signals. We found good production and complementation with an interleukin-12 secretion signal linked to the SmBiT-ACE2 fusion protein while an IgK secretory leader sequence worked best at the N-terminus of the RBD-LgBiT protein. Transfection of these constructs into 293T cells showed strong expression in cell lysates and secretion into supernatants, as confirmed by immunoblot analyses (Fig. 1C). We then carried out luciferase assays using either cell lysates or supernatants from 293T cells co-transfected with RBD and ACE2 NanoBiT fusion constructs in both orientations (Fig. 1D). As can be seen, SmBiT-ACE2 and either RBD-LgBiT or LgBiT-RBD (>105 RLU vs ~104 RLU in Fig. 1C) produced strong luminescent signals compared to transfection of individual components.
Supernatants from the co-transfected cells also produced strong luminescence in the assay mirroring our findings with the cell lysates (Fig. 1D). The large dynamic range of the assay makes it amenable to high-throughput screening and indeed bioluminescent signals could even be observed using an IVIS imaging system or with the naked eye (Supplementary Fig. 2B). We validated the specificity of the interactions we were detecting in two ways. First we co-transfected SmBiT-ACE2 or LgBiT-RBD with LgBiT- YAP15 or SmBiT-14-3-3, respectively. While the YAP15 and 14-3-3 constructs have been previously demonstrated to interact and rescue the NanoBiT structure11, these proteins are not known to interact with ACE2 and RBD. As predicted, LgBiT-Yap15 and SmBiT-14-3-3 proteins did not complement SmBiT-ACE2 or LgBit-RBD respectively (Fig 1E ). Second we evaluated the impact of recombinant soluble ACE2 protein (rACE2), a SARS-CoV-2 entry inhibitor under clinical investigation as an antiviral22, and recombinant RBD (rRBD) on the SARS-CoV-2 NanoBiT. For these experiments, we transfected 293T cells independently with either the RBD-LgBiT or SmBiT-ACE2 constructs to obtain lysates or culture supernatants containing either RBD-LgBiT or SmBiT-ACE2. In the absence of added recombinant protein, the combined lysates or supernatants produced robust signals (Fig. 1F). However, upon pre-incubation of RBD-LgBiT with rACE2 prior to adding the SmBiT-ACE2 containing lysate, we observed dose dependent reduction in luminescent signal (Fig. 1F). Similarly, pre-incubation of SmBiT- ACE2 with recombinant RBD resulted in the loss of luminescence in the biosensor reporter assay (Fig. 1G). These results serve as proof-of-principle that our biosensor can identify molecules which disrupt the ACE2-RBD interaction. We also constructed a SARS-CoV-1 NanoBiT biosensor. Earlier work suggested that the SARS-CoV-2 RBD binds with higher affinity to ACE2 than SARS-CoV-1 RBD, potentially contributing to the virus’ enhanced transmissibility8 while others have shown that the RBDs from these two viruses have comparable affinities7, 23. In our hands, the SARS-CoV-1 NanoBiT produced only a modestly reduced signal relative to the SARS-CoV-2 biosensor (Fig. 1H and Supplemental Fig. 4) supporting the idea that two virus RBDs have comparable affinity for ACE2.
The SARS-CoV-2 Biosensor is Sensitive to Neutralizing Antibodies
Monoclonal antibodies targeting RBD are under consideration as SARS-CoV-2 therapeutics24. We screened 13 different commercially available monoclonal SARS-CoV-2 Spike RBD antibodies with the SARS-CoV-2 biosensor (Fig. 2A). Seven of these antibodies (# 4,5,6,7,8,9,10) are reported to not only bind RBD but also neutralize infection of cells with an S pseudotyped lentivirus. Interestingly these seven monoclonal antibodies were the most effective at blocking RBD-ACE2 interactions measured with the SARS-CoV-2 biosensor. We applied the antibody collection to the SARS-CoV-1 biosensor and found that while most SARS-CoV-2 antibodies did not cross-react, antibodies #7 and 8 showed some ability to disrupt RBD-ACE2 interactions for both virus strains. Non-specific mouse IgG and monoclonal antibody #1, which binds to the S2 subdomain of the Spike protein, did not disrupt the signal generated by the SARS- CoV-2 biosensor supporting the specificity of the signals were observed. We tested our biosensors with serum from two patients recovered from SARS-CoV-2 infections at the Ottawa Hospital and pooled serum from three healthy volunteers. In these experiments we compared our SARS-CoV-2 biosensor to a widely used, commercially available ELISA kit that is designed to act as surrogate for virus neutralization. For the biosensor experiments, SARS-CoV-2 RBD-LgBiT was co-incubated with sera for 25 min, followed by the addition of SmBiT-ACE2 for an additional five minutes. At this point, substrate was added and luminescence measured. The biosensor was able to distinguish seroconverters from healthy donors as both convalescent patients’ sera significantly reduce the biosensor signal (Fig. 2B- right panel). Interestingly, sera from convalescent SARS-CoV-2 patients failed to disrupt the SARS-CoV-1 RBD-ACE2 interaction (Fig. 2B- left panel), suggesting a lack of cross-reactivity in these patients’ neutralizing antibody response. The signal from our biosensor compared well with the results produced by a receptor-ligand binding ELISA from GenScript (Fig. 2C). It is worth noting, however, the NanoBiT assay is a more rapid and simplified single tube reaction.
SARS-CoV-2 genome sequencing has revealed the emergence of RBD mutations in global strains. We investigated the influence of six emerging RBD mutations found in SARS-CoV-2 genome sequences worldwide on the ACE2-RBD interaction: V367F (France and Hong Kong/China), N354D (China), A435S (Finland), F342L (England), K458R and V483R (United States)25 (Fig. 2D). The biosensor assay revealed that these SARS-CoV-2 variants displayed variable binding to ACE2 (Fig. 2E- F). Interestingly, the V367F mutant displayed over 3-fold enhanced interaction with ACE2, while the F342L mutation decreased reporter activity 2-fold. The enhanced affinity of V367F RBD mutation to ACE2 is consistent with a recent study describing enhanced viral entry in HEK293T-ACE2/TMPRSS2 cells with lentivirus pseudotyped with V367F Spike compared to wildtype Spike9. Similarly, these mutations also have the potential to impact the efficacy of RBD-targeted monoclonal antibodies and vaccination strategies. We analyzed the cross-reactivity of two SARS-CoV-2 RBD- targeted monoclonal antibodies towards the different RBD variants using the biosensor assay (Fig. 2G-H). Our results demonstrated that both monoclonal antibodies tested could effectively block all the mutants’ interactions with ACE2 – highlighting that monoclonal RBD antibodies can work effectively against SARS-CoV-2 strains encoding the different RBD variants. As described below, we determined that the RBD used in our biosensor, when produced from human 293T cells, is glycosylated at the two asparagine residues sites previously identified using a site-specific mass spectrometric approach. Taken together, our results suggest that, N-linked glycans are ineffective in blocking the recognition of RBD by a spectrum of neutralizing antibodies and polyclonal patient serum. This prompted us to determine if modifications at these two sites could play a role in RBD binding to ACE2.
N-linked glycosylation of RBD is required for ACE2-RBD interaction
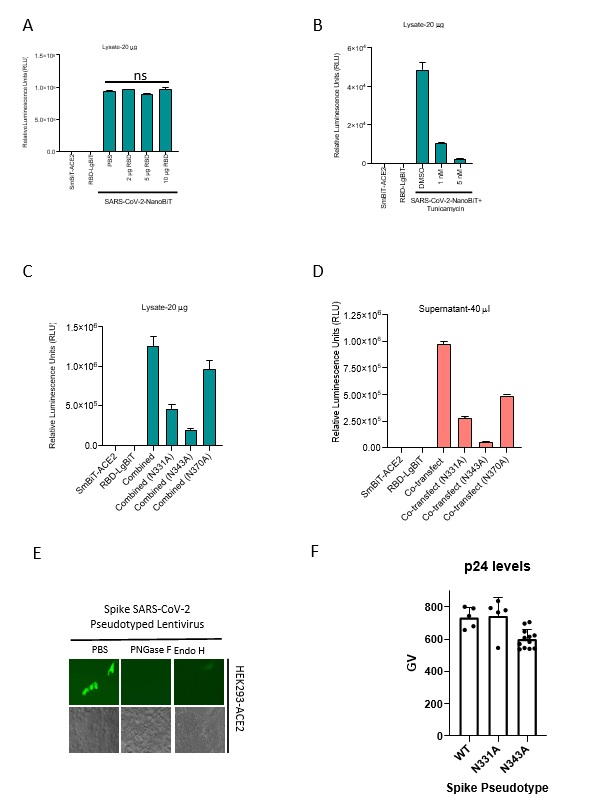
We found that bacterially produced recombinant SARS-CoV-2 RBD was not able to block SmBiT-ACE2’s interaction with SARS-CoV-2 RBD-LgBiT (Supplementary Fig. 5A), in contrast to our results with recombinant RBD produced in mammalian cells (Fig. 1G). An important distinction with bacterial expression systems is their inability to produce mammalian-type glycosylation, suggesting a potential role for protein glycosylation in the ACE2-RBD interaction. As discussed earlier, a recent study demonstrated that the spike protein contains 22 N-linked glycosylation sites6 including two in the RBD at asparagine residues 331 and 343. To evaluate the relevance of N- linked glycosylation of RBD on the ACE2-RBD interaction, we pre-treated RBD-LgBiT or SmBiT-ACE2 containing lysates with peptide:N-glycosidase F (PNGase F) or endoglycosidase H (Endo H), enzymes, which cleave N-linked oligosaccharides, and subsequently interrogated their ability to interact with their complementary partner using the biosensor assay. Interestingly, RBD-LgBiT’s treatment with either enzyme significantly impaired its interaction with the ACE2 ectodomain, whereas treatment of SmBiT-ACE2 had no effect (Fig. 3A). We next performed the biosensor assay using RBD-LgBiT cell lysates derived from 293T cells treated with tunicamycin, an inhibitor of N-linked glycosylation in eukaryotic cells. Immunoblot analyses demonstrated that RBD-LgBiT transfected cells which were treated with tunicamycin produced RBD-LgBiT with an apparent lower molecular weight – suggesting a loss of glycosylation (Fig. 3B). Tunicamycin treatment resulted in significantly reduced biosensor activity in lysates from cells co-transfected with RBD-LgBiT and SmBiT-ACE2 (Fig. 3C, Supplemental Fig. 5B). Taken together, this data suggests that N-linked glycosylation of the SARS-CoV-2 S protein RBD is necessary for its interaction with ACE2. To more directly determine the importance of RBD glycosylation, we substituted alanine residues for aspargines at positions 331 and 343. Immunoblot analyses of the point mutations in RBD-LgBiT revealed that N331A and N343A mutations resulted in decreased molecular weights, consistent with the two sites being functional glycosylation sites (Fig. 3D) and paralleled the changes in mobility of RBD produced from tunicamycin treated cells. We evaluated the impact of mutating these two glycosylation sites on ACE2-RBD interactions using our biosensor assay (Fig. 3D-E; Supplemental Fig. 5C-D). Both the N331A and N343A mutations significantly impacted complementation between ACE2 and RBD fusion proteins. To further validate the importance of N-linked glycosylation to SARS-CoV-2 infectivity using an orthogonal approach, we also used Endo H and PNGase F treatment on S protein pseudotyped lentiviruses. Consistent with the biosensor data (Fig. 3A-E), enzymatic removal of N-linked glycosylation abrogated the infectivity of the S pseudotyped lentivirus (Fig. 3F). We then used site-directed mutagenesis to create full-length Spike mutants (N331A and N340A) and used these to create S pseudotyped lentiviruses. Consistent with our SARS-CoV-2-NanoBiT data (Fig. 3E), both mutations produced significant decreases in S pseudotyped lentivirus infectivity (Fig. 3G, Supplemental Fig. 5E-F). Overall, these data provide the first direct evidence that SARS-CoV-2 S depends on N-linked glycosylation of RBD to mediate its interaction with the ACE2 ectodomain.
SARS-CoV-2-NanoBiT identifies lectins as antiviral therapeutic candidates
SARS-CoV-2 S is glycosylated with oligomannose- and complex-type glycans 6. We sought to examine the therapeutic potential of targeting these N-linked glycans by testing mannose-binding plant lectins for anti-viral effects. We screened lectins from Canavalia ensiformis (jack bean), Pisum sativum (pea), Galanthus nivalis (snow drop),
Datura stramonium (jimson weed/thorn apple), and Lens culinaris (lentil) for their ability to disrupt the SARS-CoV-2 RBD-ACE2 interaction using our biosensor (Fig. 3H). Our results demonstrate a diverse range of antiviral effects. While the lentil lectin displayed no significant inhibition of the interaction across the tested concentration range (8-1000 ng/mL), other lectins showed some efficacy, with the jack bean lectin (Concanavalin A or ConA) demonstrating the strongest impact. The antiviral effects for the top two lectin candidates (Pisum sativum and ConA) were validated for their ability to inhibit Spike pseudotyped lentivirus at 100 ng/mL (Fig. 3I). Consistent with the CoV-NanoBiT data, both lectins inhibited pseudovirion entry, with ConA showing over a 900-fold decrease while the pea lectin showed only an 8-fold decrease. ConA’s antiviral effects are consistent with previous work suggesting mannose-binding lectins can inhibit SARS- CoV infection 26. Collectively, our work suggests targeting the glycosylation of Spike represents a viable therapeutic target that warrants further investigation.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}