Malaria and HIV/AIDS are responsible for high morbidity and mortality rates in endemic regions of Sub-Saharan Africa, where they largely overlap in their geographical distribution and remain major health concerns. Even though the infectious agents of the two pathologies share blood as biotope, their epidemiology differs a lot and depends upon the geographic region. Nevertheless, fever is the most frequent and prominent symptom for both diseases, but it is non-specific, since it can result from the release of hemozoin in malaria infections, or from opportunistic pathogens proliferation in HIV/AIDS. It is defined as a condition of the human body with temperature higher than 38°C [1]. As previously said, fever is the main symptom of malaria, but also a common manifestation of many infectious diseases. Malaria infection becomes a disease when parasitaemia above a certain threshold leads to the production of signs and symptoms [2–3]. However, in studying the specificity of highland malaria in Cameroon, it was discovered that these malaria-related symptoms vary with altitude [4]. Management of Plasmodium and HIV co-infection should therefore take into account the malaria epidemiological level of the locality.
In holoendemic areas of malaria, plasmodial infection could have effects on HIV acquisition, as well as its progression and response to therapy [5–7]. Moreover, HIV infected individuals when bearing malaria parasites, have significant increase in viral load and decrease in T-lymphocytes CD4 count [8]. The phenomenon is even more severe in HIV infected adults [9]. Furthermore, the plasmatic viral load of HIV increases during the course of malarial episode in absence of treatment [5]. Conversely, levels of malaria parasiteamia are positively correlated to the intensity of the immunity deficit [10]. HIV/AIDS can increase the adverse effects of malaria, including anaemia and placental Plasmodium infection [11].. HIV infection may also leads to an increase of the Plasmodium parasitaemia, along with the frequency of clinical malaria; probably by suppressing the anti-Plasmodium immunity of the infected subjects [12–16]. In fact, viral infection induces cellular destructions, which lead to a decrease in T-CD4 lymphocytes, as well as in their function within the cellular mediated immunity response [17]. This disruption of the acquired immune response also affects incidence, frequency, severity and even the efficacy of anti-malarial drugs [17–19]. It is important to note that in hyperendemic zones of malaria, HIV leads to an increase of malaria parasitaemia, thus inducing an increase of the infectious reservoir within the human population and the subsequent transmission rate of malaria [05], though a trend of increase in transmission-blocking immunity was observed with rising gametocytaemia in the general population [20]. These observations raise the question of possible interactions between the two pathogens in holoendemic areas, already discussed but still unclear [21]. However, in hypoendemic zones like highlands where malaria transmission and incidence are low, and populations are instead prone to epidemics, interactions between Plasmodium and HIV in case of co-infection have not yet been studied.
There are concerns that where HIV prevalence is high, co-infection of HIV and Plasmodium does exist [22], and the use of antiretroviral drugs (especially protease inhibitors), in some cases inhibits liver stage parasites at clinically relevant concentration [23], and in other cases can affect the efficacy of the newly introduced artemisinin-based combination therapy (ACT) [24]. The impediment in this situation is that, simultaneous administration of anti-malarial with anti-retroviral treatments can lead to high level of toxicity [25–26]. Treatment of HIV and Plasmodium co-infection is therefore a critical issue, and should be considered as medical emergency.
Liver involvement in Plasmodium infection and the subsequent function-abnormalities or dysfunctions are common in severe malaria and may manifest as elevated liver enzymes or hyperbilirubinemia [27–28]. In fact, infection of liver cells can cause organ congestion, sinusoidal blockade and cellular inflammation. These changes in hepatocytes can lead to the leakage of parenchymal transaminases and membranous phosphatase enzymes of the liver into the blood circulation [29]. It has been shown that if malaria is neglected or not treated properly, there will be an increase in the blood level of transaminases which will cause damage to the liver cells [30]. As mentioned above, hepatocellular dysfunction is frequent in malaria, but that qualifying as malarial hepatopathy is not common. It appears mostly in case of severe malaria and is more likely to occur in presence of other complications [31]; hence coinfection with HIV could favour its development. Moreover, individual-specific factors may confer more or less susceptibility to hepatocyte injury [32]. The importance of proving whether or not Plasmodium infection induces greater hepatocellular disease in immunodeficient people is of great consequence, because if such were the case, these patients should be treated for hepatitis as well as malaria.
Following the release of metabolic wastes present in the digestive vacuole of Plasmodium (from the rupture of infected erythrocytes) into blood circulation during malaria attack, hemozoin is engulfed and accumulated within circulating monocytes and tissue macrophages (reticulo-endothelial system). It mediates the release of endogenous pyrogens that altered several functions of the immune system; especially thermoregulation [33]. This hemozoin accumulation is associated with some malarial symptoms, mainly fever. In fact, high levels of hemozoin within immune cells have been shown to correlate with disease severity [34–35]. However, the physiopathology of the host-parasite interactions and particularly the mechanism of fever induction in malaria have not been clarified yet. The slight progress made until recently, about the potential mechanism of malaria-induced fever reports on a novel mechanism that the host uses to recognise Plasmodium DNA via a Toll-like receptor (TLR–9). It has elucidated an important step in the long-time discussion on why and how does malaria cause fever, by showing that hemozoin internalises plasmodial DNA and present it to TLR9. Plasmodial DNA then intracellularly interacts with TLR9, initiating signal transduction leading to the release of proinflammatory cytokines. These cytokines induce production of prostaglandins which subsequently leads to induction of fever. Hemozoin is therefore involved in malaria fever but is toll-free [36]. This discovery corrects previous findings claiming that hemozoin was a direct TLR9 stimulus, and that stimulation of TLR–9 by hemozoin pigment could be the key step for inducing fever in human host during malaria attack [37]. Some authors have pointed out a production of prostaglandins that are pyrogenic and immunosuppressive, by both the human host cells and Plasmodium themselves [38]. It is likely that these important mechanisms of malaria physiopathology may also apply to opportunistic infectious diseases in people living with HIV (PLHIV). Anyhow, more knowledge on the fever physiopathology is needed in order to discriminate the sources of fever in co-infected patients, and develop intervention strategies towards reducing the disease burden.
Fever episodes are usually attributed to malaria in endemic zones of Sub-Saharan Africa, because the case definition of symptomatic malaria generally used is fever associated with a specific parasite density threshold [30–31, 39–40]. For this reason, it has been recommended for quite a long time that for all febrile episodes, a presumptive treatment should be administered [41]. The approach was of value to clinicians, but not to epidemiologists. In fact, it is not clear so far, whether the presumptive treatment of fever episodes as malaria cases is an advantage or a hindrance for malaria control [42]. Recently, with achievement of the rapid diagnostic test that enable quick and easier examination of blood, WHO moved away from presumptive treatment to one that advocates parasitological diagnosis whenever possible [43]. The aim of this shift of policy was to avoid treating large numbers of non-malarial fevers, which represent a non-negligible fraction in holoendemic areas, in contrast to highlands. The potential benefits of parasitological diagnosis will depend mainly upon the prevalence of Plasmodium infection among patients who report fever (Pf, knowing that these malarial fevers are always associated to other manifestations [44–45]. Unfortunately, this policy is not practically implemented in most places. Yet, treatment of fever in our hyperendemic region ought to be done with extreme caution, since several infectious diseases also generate fever; especially opportunistic diseases in people living with HIV infection (PLHIV). In fact, a bright confusion surrounds the definition of fever and its use in clinical diagnosis of malaria [46–49]. For some authors, fever or any recent febrile episode strongly predicts a malarial infection [22]; while for others, it is more likely associated to an infectious disease [50]. Nevertheless, it is intuitively obvious that PLHIV in malaria hyperendemic zones will present an increased risk of developing malaria, since their immune system is deficient [11–14]. However, very little is known about hypoendemic areas.
Since the prevalence of malaria among PLHIV was not known in the hypoendemic area of Western-Cameroon highlands, it was important to make a distinction between Plasmodium-induced fever and the others, for a better treatment policy of these peculiar patients. This study aimed principally at evaluating the malaria-attributable fraction of fever episodes (MAFE) among HIV-seropositive patients in the Western-Cameroon highland.
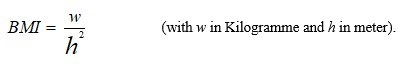{kind=link}
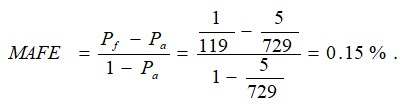{kind=link}
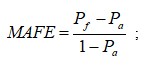{kind=link}