ERRα, VDR and RXR basal expression in tested breast cancer cells
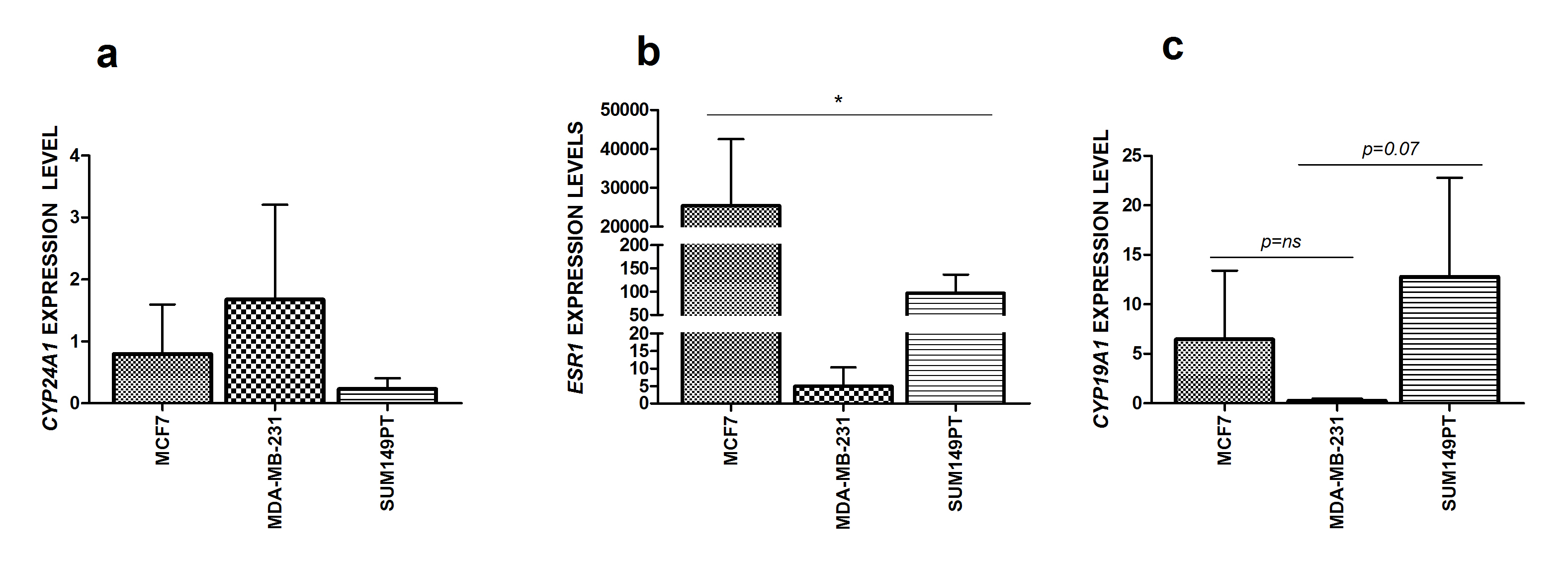
We first evaluated the expression levels of ERRα, VDR and RXR transcripts in the MCF7, MDA-MB-231 and SUM149PT cells based on our hypothesis that these biomarkers may affect the response to calcitriol. The real time evaluations, performed by using bona fide normal MCF 10A cells as a reference, showed that a higher ERRα mRNA level was found in SUM149PT cells compared to MCF7 and MDA-MB-231 (though it did not reach the significant p value, p = 0.08); (Fig. 1a). The VDR transcript levels were lower in the MDA-MB-231 (p = 0.004) and MCF7 (p = 0.05) cells than in the SUM149PT (Fig. 1b) cell line; while no significant difference was found among the three cell lines in the basal RXRα mRNA levels (Fig. 1c). Since our focus was on the calcitriol degrading enzyme and estrogen signaling, we determined the basal expression levels of the CYP24A1, ESR1 and CYP19A1 transcripts (Supplementary Figure S2). Collectively, these data pointed out that the SUM149PT cell line showed the highest expression levels of both the VDR and ERRα transcript, while there was no significant difference regarding the CYP24A1 and CYP19A1 expression levels. As expected, unlike MCF7, which is an ER + luminal A breast cancer model, both MDA-MB-231 and SUM149PT displayed barely detectable levels of ESR1 since they represent triple negative breast cancer models. [32].
Genomic effects of the calcitriol/VDR axis: focus on the calcitriol degradation enzyme, CYP24A1 and the estrogen pathway
Next, to explore the genomic action of VDR, we challenged the cells with 100 nM calcitriol, which is the concentration generally used to study the effects of VDR activation [33]. We found that CYP24A1 transcript expression rapidly increased after 4 h of calcitriol treatment in SUM149PT cells (> 500 fold and > 50 fold over the vehicle-treated cells; p = 0.004) (Fig. 2a) and further increased after 24 h of treatment (> 10000 fold and > 1000 fold over the vehicle-treated cells; p = 0.004) in MCF7 cells (Fig. 2b). CYP24A1 transcript expression increased to a lesser extent in MDA-MB-231 cells than in the SUM149PT and MCF7 cell lines. It was about 2 fold greater than in the vehicle-treated cells (p = 0.01) after 4 h of calcitriol (Fig. 2a), and up to 3 fold greater than in the vehicle-treated cells (p = 0.22) after 24 h of treatment (Fig. 2b). Given that Santoz-Martinez et al. reported that 100 nM calcitriol induced the expression of a functional ERα in the MDA-MB-231cell line [17], and we hypothesized a functional interaction between VDR and ERRα that may activate estrogen signaling, we also determined the effect of calcitriol on the expression of ESR1 and CYP19A1 transcripts. We found that 100 nM calcitriol induced a time-dependent stimulation of ESR1. ESR1 transcript levels were more than 1 fold higher in SUM149PT cells than in the vehicle-treated cells (p = 0.03) by 4 h, and more than 3 fold higher than in the vehicle-treated cells (p = 0.02) (>) (Fig. 2c) after 24 h of calcitriol treatment (>) (Fig. 2d), while a transient stimulation of ESR1 transcript occurred only after 4 h (p = 0.01) in MDA-MB-231 cells (Fig. 2c). Calcitriol did not significantly modulate ESR1 gene expression in MCF7 cells (Fig. 2c-d). A slight but significant induction of CYP19A1 transcription (p = 0.03) occurred by 4 h (Fig. 2e) in the MDA-MB-231 cells, but it was no longer detectable after 24 h of treatment (Fig. 2f). Aromatase transcript levels increased in the MCF7 cells (> 1 fold higher than in the vehicle-treated cells p = 0.04) and to a much greater extent in the SUM149PT cells by 24 h (> 14 fold higher than in the vehicle-treated cells p = 0.007) (Fig. 2f). Collectively, our findings demonstrated that in the SUM149PT cell line calcitriol strongly induced the expression of its degrading enzyme (CYP24A1) as well as of key estrogen signaling biomarkers. We thus chose the SUM149PT cell line to assess the role of ERRα in the biological behavior of VDR in a representative model of triple-negative, inflammatory breast cancer falling within the most aggressive Basal-like subtype of BC (BLBC), and the MCF7 cell line for the same purpose in a Luminal A breast cancer model that is less invasive and aggressive.
ERRα loss of function abrogated VDR genomic action on CYP24A1, ESR1 and CYP19A1, but activated that on KDM1A
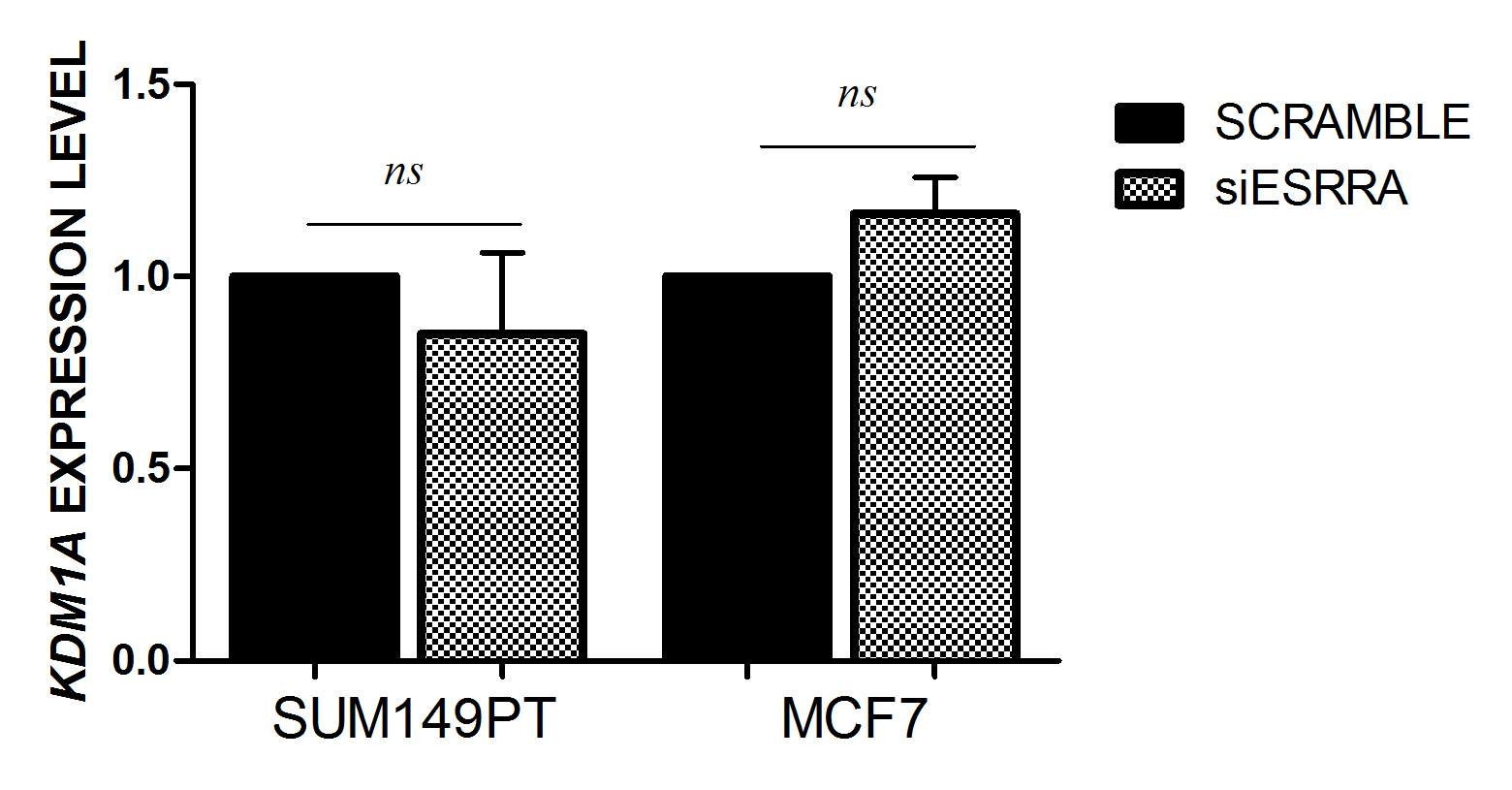
To investigate the biological function of ERRα on calcitriol/VDR genomic action, MCF7 and SUM149PT cell lines were treated with 100 nm calcitriol, after the cells had been transfected with siRNA targeting ERRα or with negative control (NC). Knockdown of ERRα restored the basal expression of CYP24A1 in both SUM149PT (p = 0.0003) and MCF7 (p = 0.01), thus completely abrogating the effect of calcitriol on its degrading enzyme (Fig. 3a). Remarkably, ESR1 expression also decreased and returned to its basal level in SUM149PT cells (p = 0.0008) (Fig. 3b), and the same happened to CYP19A1 transcript in both MCF7 (p = 0.009) and SUM149PT cell lines (p = 0.03) (Fig. 3c). These results suggest that ERRα was a crucial coactivator for the VDR transcription complex to carry on a program leading to calcitriol degradation and activation of estrogen signaling. Of note is that this phenomenon occurred to a higher extent in the basal-like model than in the luminal A model. Recently, Battaglia et al. reported that LSD-1 mediated the epigenetic corruption of vitamin D signaling in prostate cancer [26], and Carnesecchi et al. reported a close interaction between ERRα and LSD1 to regulate each other, mostly in cancer cell invasive behavior [27, 28]. In particular, LSD-1 was involved in the maintenance of ERRα protein stability, while the ERRα protein induced LSD-1 to erase repressive marks in vitro, thereby promoting the transcriptional activation of genes involved in the invasion of the extracellular matrix. Hence, we explored the effect of ERRα silencing on KDM1A expression upon calcitriol treatment to gain insights into the functional interaction of ERRα and KDM1A in VDR signaling in BC. Interestingly, ERRα silencing did not alter KDM1A expression in either cell line (Supplementary Figure S3) while calcitriol treatment significantly upregulated the mRNA expression of KDM1A only in transfected SUM149PT cells (Fig. 3d).
Effect of ERRα knockdown on cell clonality, calcitriol cytotoxicity and underlying mechanisms
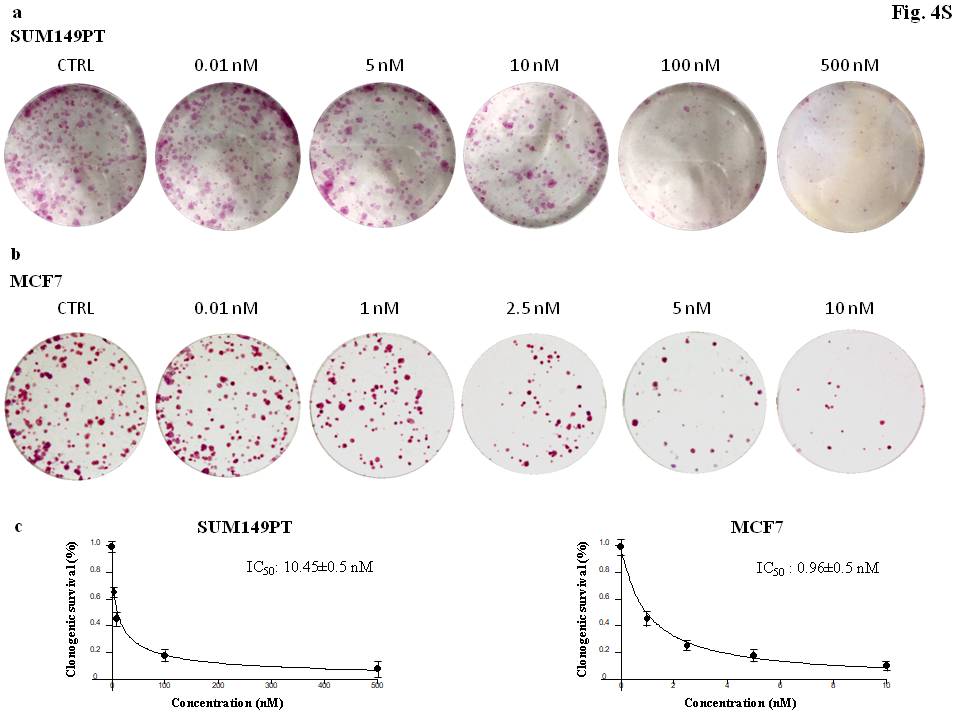
To assess whether ERRα influenced tumor cell proliferation and sensitivity to calcitriol, we first tested the effect of single treatments –either calcitriol or ERRα knockdown– on cell clonality and then we tested the effect of the combined treatment. The results of colony formation assays indicated that i) calcitriol induced a concentration-dependent reduction of the numbers and size of colonies in both cell lines, with MCF7 cells being the most sensitive to calcitriol (data reported as supplementary material (Supplementary Figure S4) ii) ERRα knockdown significantly reduced colony formation in both cell lines (Fig. 4a-4d) iii) by contrast, ERRα silencing abrogated calcitriol cytotoxicity in SUM149PT cells and strongly reduced it in the MCF7 cell line. Calcitriol reduced colony formation in MCF7 much less than in non-silenced cells (si-NC + calcitriol) (Fig. 4a-4d). Since estrogens preferentially induce cyclin D1 to trigger breast cancer proliferation while p21 is transcriptionally regulated by ERRα to remove constraints in tumor progression [34], we evaluated the function of ERRα in the expression of these targets and in VDR protein expression to explore the potential regulatory mechanism of sensitivity to calcitriol. We found that calcitriol induced an increase in VDR protein expression in both cell lines in ERRα-silenced cells and in ERRα-expressing cells (transfected with si-NC), meaning that VDR activation occurred [35] irrespective of ERRα expression. However, by comparison, calcitriol reduced cyclin D1 expression in si-NC-MCF7 (control) cells to a much greater extent than in si-ERRα-MCF7 cells, while no effect was observed on p21 expression in both. By contrast, calcitriol increased p21 expression in si-NC-SUM149PT cells much more than in si-ERRα-SUM149PT cells, while no variation was found for cyclin D1 expression (Fig. 4c-4f). Accordingly, the data on gene expression showed that p21 was regulated by ERRα in SUM149PT cells, as ERRα silencing significantly upregulated p21 in the SUM149PT cell line and not in MCF7 cells (Fig. 4g). Target modulation was reflected at the level of cell cycle progression. Calcitriol caused G0/G1 phase cell cycle arrest in both SUM149PT and MCF7 cells while combination with ERRα-targeting treatment abrogated the effect of calcitriol on the cell cycle in both cell lines (Fig. 4b-4e). Collectively, the results indicated that ERRα supported proliferation in both cancer models. Our findings suggested that, although a preferential involvement of ERRα conveyed sensitivity to calcitriol in SUM149PT cells while ERα did so in MCF7 cells, ERRα was crucial for the tumor-suppressive ability of calcitriol in both tumor models, which is in line with the ability of ERRα and ERα to interfere and collaborate each other as demonstrated by their co-regulation of several common target genes [36].
VDR and ERRα cellular localization in MCF7 and SUM149PT cells after calcitriol treatment
To further address a VDR and ERRα interaction, we performed immunofluorescence analysis to visualize the cellular distribution of VDR and ERRα in calcitriol-treated cells versus vehicle-treated cells. As shown in Fig. 5a, time-dependent nuclear accumulation of VDR and ERRα was observed in SUM149PT cells, in which both nuclear receptors basically co-localized after treatment with calcitriol. The MCF7 cell line showed a higher basal ERRα expression in the nucleus, unlike VDR. Upon calcitriol treatment both ERRα and VDR increased in the nucleus (Fig. 5b). Consistent with data we reported before, immunofluorescence results suggest that VDR and ERRα interact and that their interaction is ligand-dependent in SUM149PT cells and ligand-enhanced in MCF7 cells. To examine whether this was a result of a direct interaction we performed a bioinformatics analysis.
Interactome analysis of VDR/ESRRA axis
An interactome analysis was set-up. Through BioGRID, we built an extended network to query VDR, ESRRA, ESR1, BRCA1 and KDM1A as main interacting protein hubs. Such a choice was based on ESRRA interacting proteins emerged by our study and by recent report showing a direct interaction between ESRRA and BRCA1 in BRCA1-mutated carriers [37], which is a setting represented by the SUM149PT cell line in our experiments, while others have demonstrated a direct interaction between ESRRA and KDM1A [28, 27]. The subnetwork, identified from the whole database (Fig. 6a), evidenced a cluster of 31 interacting proteins and was further analyzed through the STRING interaction database. This further analysis allowed to better explore the BioGRID subnetwork, highlighting the specific types of interactions connecting the nodes (Fig. 6b). Of note, PPARGC1A played a central role in the directed network, connecting VDR and ESRRA.
From biological network to pathway cross-talk
The impact of our in vitro results was studied in the TCGA-BRCA cohort. Cases were selected according to molecular subtypes in order to reflect the setting of cell lines, namely Basal-like (SUM149PT) and Luminal A (MCF7). After the selection step, the in silico cohort included 567 patients with Luminal A BC and 194 patients with basal-like BC.
In order to assess the pathway activity related to the VDR/ESRRA axis, the FPKM values of the KDM1A, BRCA1 and PPARGC1A genes were also included in the crosstalk analysis, given the roles they proved to have in our in vitro experiments and according to our interactome results. The StarBioTrek package was used because it is more informative than enrichment analysis in providing information on pathways and their relative cross-talk integrating networks and gene expression data. We found “control of gene expression by vitamin d receptor”/“regulation of pgc-1a” and “pelp1 modulation of estrogen receptor activity”/“control of gene expression by vitamin d receptor” crosstalks with AUC values of 0.55 and 0.52, respectively, using Biocarta pathway data integrated with PHint network information. We thus tried to identify the biological role of ESRRA by dichotomizing the basal-like subset for its expression. Interestingly, we detected the same crosstalks as in the previous comparison with AUC values of 0.67 and 0.66, respectively in the ESRRA overexpressing group. Such a result is promising because ESRRA stratification is able to biologically discriminate basal cases with more elevated AUC values than a basal-like versus Luminal A group analysis.
Translational significance of ESSRA/VDR axis and survival in TCGA dataset
Literature data and our in vitro and in silico results left the prognostic value of the VDR-CYP24A1-ESRRA axis open to question. Overall survival data of basal-like patients were downloaded and the patients were stratified into two groups according to whether VDR-CYP24A1-ESRRA simultaneous overexpression was present or not. The Kaplan-Meier curves (Fig. 6c) and log-rank test showed that patients overexpressing VDR-CYP24A1-ESRRA genes had a significantly worse survival than the other group (p-value = 0.017), clearly indicating a prognostic value of such a biomarker signature for basal-like breast cancer.
{kind=link}
{kind=link}
{kind=link}
{kind=link}