Chromosomal aberrations detected in 2D cell culture
Genomic profiling of trypsinized SK-N-BE(2) cells grown traditionally in monolayer culture revealed the presence of ten SCAs detected by HD-SNPa, as previously reported [42]: three SCAs typically observed in NB and seven SCAs classed as atypical for being less frequently present in NB. This cell line also showed a numeric chromosome alteration (NCA). As already described, SK-N-BE(2) presented genome amplification in 2p24.3 involving the MYCN, MYCNUT, MYCNOS and GACAT3 genes [42,43]. A chromothripsis-like phenomenon was also detected at chromosome 21 involving sixteen fragments. Representations of genetic findings are shown in Figures 1, 2 and 3.
For the SH-SY5Y cell line grown in 2D culture, as previously reported[44], we found five SCAs and two atypical SCAs. We also detected a NCA and two large copy neutral loss of heterozygosity (CNLOH) (Fig. 1). Several focal segmental chromosome alterations (FSCA, <2 Mb) and small CNLOH were also observed in both cell lines.
For SW10, we detected no chromosome aberrations.
VN-KO with SK-N-BE(2) cells revealed SCAs similar to those of stiff hydrogels with long culture time
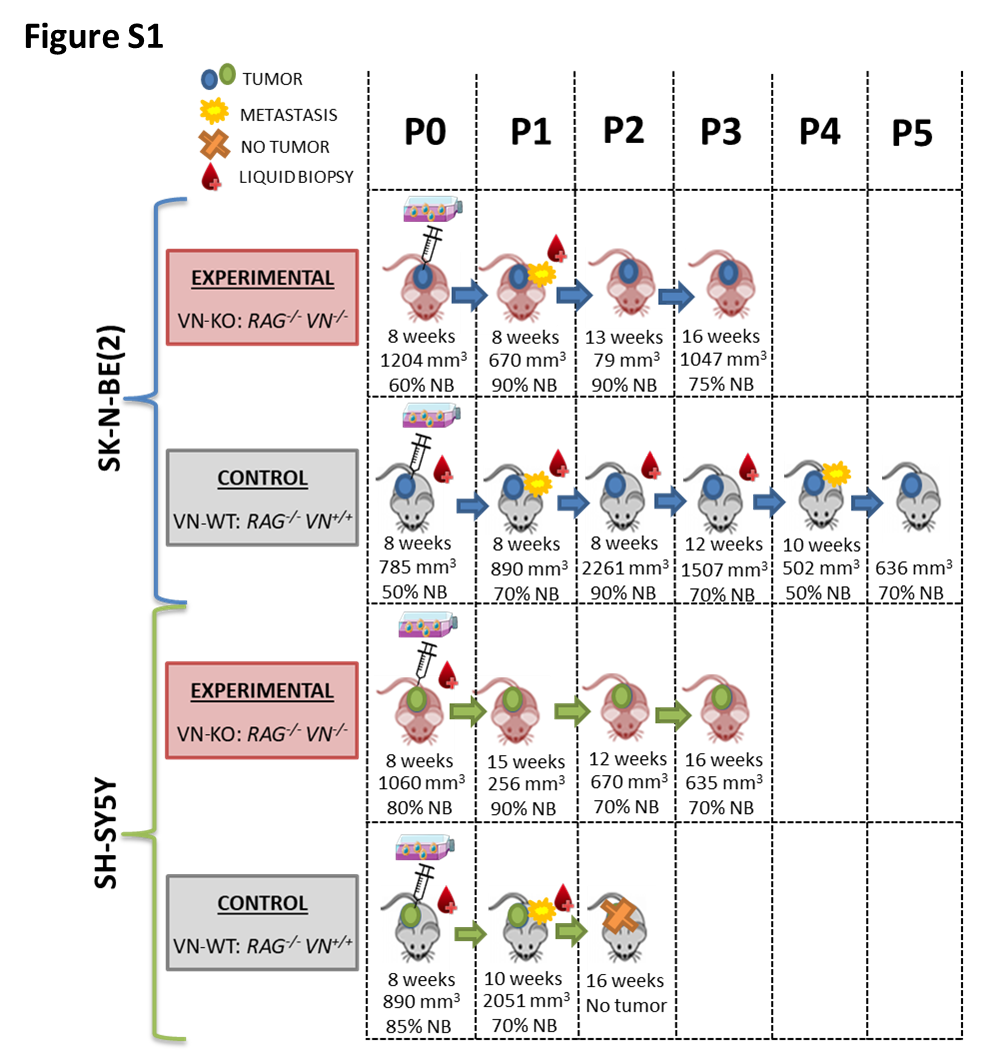
A description of each mice model (VN-KO and VN-WT) longer filum is shown in Figure S1.
P0 and P1 tumors grown in VN-KO mice showed, besides the 1p-(pter-21.3, 96.2 Mb) present in 2D SK-N-BE(2) cell line, a 1p-(21.3-12, 22Mb) and a typical +1q(21.1-qter, 105 Mb), both also present in some hydrogels. Remarkably, the first fragment contained coding genes for ECM proteins like COL11A1 and for nervous system development such as NTNG1 and NGF, and the second one contained genes with important roles in cytoskeleton, like actin-related ones (ARPC5 and ACTN2), laminins (LAMB3, LAMC1, LAMC2), and integrin α10 (ITGA10). P2 and P3 tumors reduced the latter SCA to +1q(21.3-ter, 97 Mb) in the vast majority of tumor cells (Figs. 1 and 2). Like cells grown for longer times and/or stiffer hydrogels, experimental tumors P0 and P1 showed a shorter +2p(pter-21, 43 Mb) than was observed in the 2D cell line culture, which in P2 and specially in P3 became even smaller, affecting only the smallest region of overlap (SRO) +2p(24.1-21, 21 Mb). In addition, P2 and P3 had two previously undetected genomic alterations, the typical 4p-(ter-15.2, 22 Mb) and an atypical +5q(22.2-ter, 68 Mb). A very high percentage of tumor cells from all VN-KO passages showed the same chromosomal aberrations on chromosome 9: four FSCAs (9p-(24.3), +9p(24.3-24.2), 9q-(21.13), 9q-(21.13-21.2)) and a SCA (+9q(21.2-21.33) of 8 Mb, all of them also present in stiff hydrogels with long culture periods (Figs. 1 and 2). Interestingly, 9p-(24.3) contains two important genes related to migration, KANK1 and DOCK8. The chromothripsis-like phenomenon was replaced by the fragment 21q-(22.13-22.2, 2.5 Mb) in all passages, as occurred in stiffest and longest-time hydrogels (Fig. 1. This fragment had a SRO with one of the altered fragments in the chromotripsis-like pattern: 21q-(22.13.22.2, 1.4 MB) (Fig. 1). The SCAs pertaining to the SK-N-BE(2) cell line in chromosomes 7, 11, 13, 19, and 20 were either absent or were only observed in proportionally few cells in tumors from VN-KO mice (Fig 1). Finally, NCA of chromosome 18 turned into a SCA in P1 to P3 including 18pq-(pter-q22.2, 67Mb) with the final fragment 18q(22.2-ter, 11 Mb) as a CNLOH (Fig. 1).
SK-N-BE(2) cells in VN-WT mice had high variability of SCAs between tumor passages and compared with VN-KO mice and in vitro 2D and 3D cultures. Briefly, all control mice passages changed the 1p-(pter-21.3, 96.2 Mb) for a CNLOH, maintaining only a deleted fragment 1p-(31.1-21.3, 19 Mb) in P1 and P2 tumors. The P0 sample showed a +1pq(p21.3-qter, 150 Mb), and P3 to P5 a shortest +1pq(p21.3-q23.2, 61 Mb) composed of the fragments +1p(21.3-12) and +1q(12-23). P3 to P5 tumors also had some new SCAs: 4q-(13.2-22.3), CNLOH of 17pq(pter-q11.2, 31.2 Mb), +17q(11.2-ter), +19p(ter-11) and 19q-(11-ter). These passages had neither the +7q(32.3-qter, 28.5 Mb) nor the chromotripsis-like of chromosome 21. Finally, P3 and P4 showed 7p-(ter-12.2), and P4 and P5 had +3q(26.1-ter) in some cells (Fig. 1).
HD-SNPa detected no differences in SCAs between SH-SY5Y cells in 2D cultures or when xenografted in VN-KO mice. Interestingly, in VN-WT mice without +2p(ter-16.3, 49 Mb) the P1 tumor failed to grow in any of the eight tentative tumor xenograft attempts (Fig. S1).
To determine whether xenografted tumors had intratumoral genetic heterogeneity, we analyzed HD-SNPa of ctDNA extracted from intracardiac blood (Fig S1). We observed high concordance between the SCAs of tumor DNA and ctDNA, except for two SCAs of VN-WT mice xenografted with SK-N-BE(2) cells, that were only present in ctDNAs (Fig. 1): i) P1 tumor had 2pq-(p16.3-qter, 196 Mb) in few clones (approximately 25%) and ii) P2 had the same +1pq(p21.3-q23.2, 61 Mb) as P3 to P5 in 15% of the clones.
SCAs of SK-N-BE(2) cells evolved with stiffness and culture time of 3D hydrogels
Regarding in vitro 3D scaffolds cultured with SK-N-BE(2) cells, the detected SCAs changed with AlgMA concentration and culture time. Moreover, hydrogels cocultured with Schwann cells showed an enhanced positive and negative selection of some SCAs.
In detail, cells showed the same SCAs when grown for under 4 weeks in scaffoldings as when cultured in 2D, except for the new 1p- fragment (21.3-12, 22Mb) which was found in a small percentage of cells when cultured in 1 and 2% AlgMA. The cell proportion of this SCA increased in long time hydrogels and in VN-KO mice samples (Figs. 1 and 2). Noticeably, in hydrogels with 1.5-2% of AlgMA and at least 4 weeks culturing, and hydrogel of 0.5% cultured for 10 weeks, the cell percentage of some SCAs previously observed in VN-KO mice gradually increased: i) +1q(12-qter, 105 Mb) and ii) FSCAs and SCA of chromosome 9. This positive selection was even higher in hydrogels cocultured with Schwann cells (Figs. 1 and 2). An unstable positive presence of +17q(11.2-ter, 49Mb) was found. Finally, greater stiffness and longer time of culture 3D-models, particularly those cocultured with Schwann cells, also showed clear negative selection of some SCAs, as occurred in VN-KO tumors: i) +2p(pter-16.3, 48 Mb) was gradually shortened to +2p(pter-21, 43 Mb), ii) SCAs of chromosomes 7, 11, 13, 19 and 20 showed a marked reduction of affected cells, iii) chromotripsis-like phenomenon of chromosome 21 had clonal decrease and was finally totally replaced by 21q-(22.13-22.2, 2.5 Mb).
In contrast, no genomic differences were detected by HD-SNPa between SH-SY5Y cells cultured in 2D and in 3D (Fig. 1).
Hierarchical cluster analysis is a good tool for identifying sample similarities using genomic data
Neighbor-joining tree clustering and an array of similarity and distance indexes based on the Jaccard coefficient (Fig. 3A) were obtained, according to presence or absence of the SCAs detected with ChAS software in each sample derived from SK-N-BE(2) cells (Fig. 3B). Mathematical analyses showed the highest similarity indexes between the genomics of 2D cultured cells and hydrogels with less stiffness and/or culture time. Separated from them were tumors from the control mice, with few similarities to stiff hydrogels and experimental tumors, especially with the lower indices in P3 to P5. In general, VN-WT tumors also had low similarity between their own passages.
The genomic profiles of cells derived from the stiffer and/or longer-cultivated hydrogels, and VN-KO tumor passages were grouped together, allowing us to clearly observe the differences with the softer and/or shorter time 3D-cultures and with VN-WT tumors. Note that tumors of different VN-KO tumor passages had better similarity rates.
Biotensegrity had impact in single nucleotide mutations
We sequenced a small customized panel (NB-mechanopanel) of specific genes, with mutations previously described in NB related to cytoskeleton remodeling and extracellular matrix changes, and other genes related to NB and cancer in general (Table S2). In SK-N-BE(2) cell line and in all samples derived from it, we identified two mutations described as p.C135F in TP53[45] and p.N755K in ATRX[46], defined as pathogenic in the COSMIC database. The p.I1590= and p.P1323L variants of COL11A1 (in 1p), and the intronic polymorphisms c.1680-9045C>G and c.54-20738C>T of DOCK8 (in 9p) were not detected in the majority of VN-KO tumors that had heterozygous deletions of both chromosomal regions (1p- and 9p-), despite their presence in the rest of the samples, including in the 2D cultured cell line, with allelic frequency of approximately 0.4. However, the variants p.G1504=, p.S1535P and the intronic c.652-6del of COL11A1, and the intronic polymorphism c.6068+5249A>T of DOCK8 were maintained with the same allelic frequency (nearly 1) in all the samples.
SH-SY5Y cell lines grown in 2D, as well as in 3D hydrogels and mice, showed the pathogenic mutation described as p.F1174L in ALK[47], as well as p.G12V in KRAS not previously reported in this cell line[48]. We did not find any new significant mutations (Table S2).
Chromosome 9 could be a hot spot for structural aberrations in HR-NB patients
We next sought to review the chromosome 9 profiles of homogeneous MNA primary tumor in HR-NB patients, due to the fact that the FSCAs and SCA of chromosome 9 were the main unusual genetic changes of the SK-N-BE(2) cells in the VN-KO tumor passages and in rigid and long-culture 3D-scaffolds. Surprisingly, we observed greater than expected presence of genomic aberrations on chromosome 9 in the 43 tumor profiles checked. Specifically, 39.53% of cases had aberrations affecting the chromosomal positions of the DOCK8 and/or KANK1 genes (Fig. 4).
{kind=link}