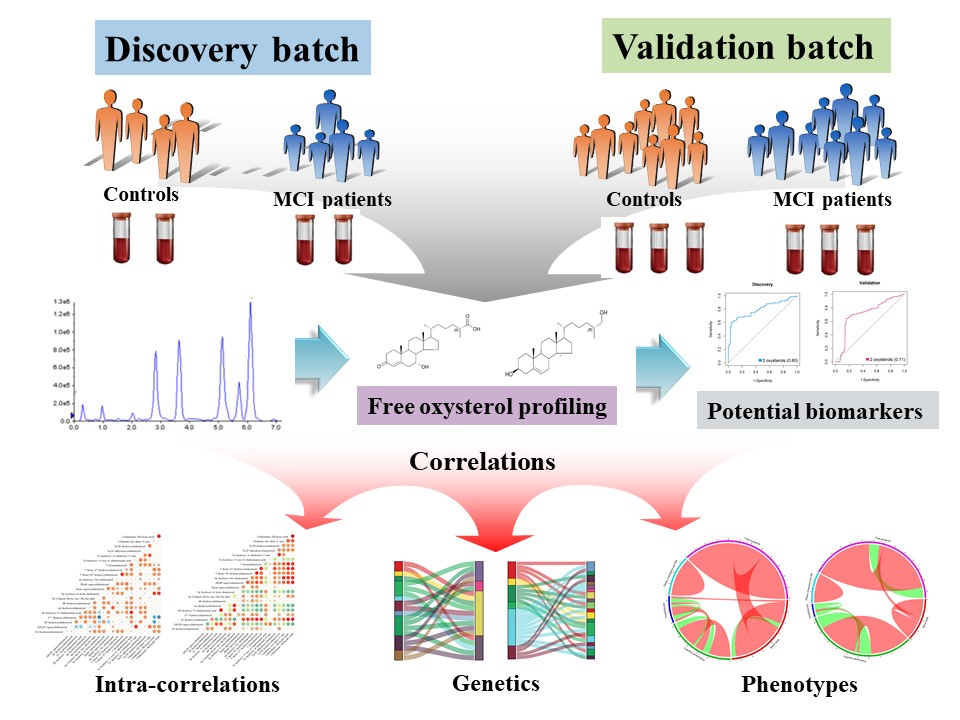
In this study, we used the ultra-performance liquid chromatography-mass spectrometry (UPLC-MS) to quantify 22 free oxysterols in serum in two groups of subjects from the EMCOA study and establish that serum free oxysterol profiles are significantly altered in MCI patients. Using ROC analyses, we identified 27-hdroxycholesterol along with its brain metabolite, 7α-hydroxy-3-oxo-4-cholestenoic acid, could distinguish MCI patients from control subjects with good performance in both the discovery batch (AUC = 0.80) and the validation batch (AUC = 0.71). We then correlated oxysterols with demographics, cognition, dietary cholesterol and fats, serum lipids and genotype and uncover how variation in specific oxysterols are linked with interindividual disparities in diet, lipids, cognition and genetic variation and how these connections are modified by cognitive status.
Accumulating research has reported significant changes in the brain concentrations and CSF levels of several oxysterols in AD or MCI patients[25, 26], which may be triggered initially by pathological changes of oxysterols in peripheral circulation. However, the measurement of blood oxysterols in MCI patients previously has gained considerable controversy and revealed little consistency[27–29]. Therefore, several unique features were incorporated into the current study in an attempt to overcome limitations of prior work and derive a more comprehensive and accurate understanding of deranged oxysterol profiles in MCI. First, rather than focusing on limited species of oxysterols, we evaluated a broader range of 22 oxysterols and their secondary metabolites. Second, the lack of verification or validation of candidate oxysterol biomarkers have been the bottlenecks of earlier research. To make the results more convincing, two independent data sets from EMCOA study were included and the initial finding in the discovery batch was further validated by a larger number of MCI patients and controls in the validation set, guaranteeing the reproducibility of our results. Third, saponification technique has been widely used in the previous research to extract “total” oxysterols without discriminating between free (biologically active) and esterified forms (largely biologically inert)[17]. In line with several most recent reports[30–32], the current study focuses on the free rather than esterified oxysterols with more sensitive method for separation and quantitative determination when taking the reasonable assumption that it is the free oxysterols that is crossing the BBB into consideration.
Here we present the discovery and validation of serum free oxysterol changes that distinguish cognitively normal participants from MCI patients. The defined two-oxysterol penal features the most common side chain oxysterol of 27-hdroxycholesterol, which mainly derives from the peripheral circulation and flows into the brain[33], and its brain-derived end metabolite, 7α-hydroxy-3-oxo-4-cholestenoic acid, to which 27-hdroxycholesterol is converted in the brain through enzymes of CYP27A1, CYP7B1 and HSD3B7 catalyzed oxidation, is then eliminated in the systemic circulation and in the CSF[12]. We also observed consistent and strong negative correlations between 7α-hydroxy-3-oxo-4-cholestenoic acid and global cognition, indicated by MoCA score, as well as processing speed performance, indicated by SDMT score. Saeed et al[34] and Crick et al[31] have demonstrated that the concentration of 7α-hydroxy-3-oxo-4-cholestenoic acid was remarkably elevated in CSF both from patients with a dysfunctional BBB and neurodegenerative disease including AD and Parkinson’s disease (PD), or amyotrophic lateral sclerosis (ALS), which may be likely to reflect increased brain accumulation of 27-hydroxycholesterol as a result of damaged BBB integrity and increased serum levels of 27-hydroxycholesterol in MCI patients. The key role of 27-hydroxycholesterol in AD pathogenesis has been strongly supported by our previous animal and cellar research, pointing to the involvement of 27-hydroxycholesterol in the pro-inflammatory molecule release[35], oxidative stress[36], lysosomal membrane permeabilization and pyroptosis[37], synaptic dysfunction[38] and Aβ production[39]. We thus discovered and validated serum 27-hydroxycholesterol and 7α-hydroxy-3-oxo-4-cholestenoic acid as diagnostic biomarkers in MCI. It is noteworthy that the currently presented serum biomarker panel of free oxysterols is especially for patients with early stages of AD development. Some studies have pointed that the brain conversion of 27-hydroxycholesterol into 7α-hydroxy-3-oxo-4-cholestenoic acid may be regarded as a protective mechanism since 7β-hydroxylated intermediates of the conversion, catalyzed by the neuronal enzyme CYP7B1, seem to be less cytotoxic[18]. The markedly reduced expression of CYP7B1 arises in the late stage of AD due to severe neuron loss and then leads to producing little or no amounts of 7α-hydroxy-3-oxo-4-cholestenoic acid[40]. Therefore, the application of this biomarker panel may be challenging in patients with long disease duration.
The relatively modest increase of other circulating oxysterol concentrations only in discovery batch suggests that these oxysterols may have limited utility as disease markers in MCI. However, many of them, especially nonenzymatical oxysterols deriving from cholesterol autoxidation, have been reported to be associated with AD progression due to their ability to stimulate inflammation[25]. Besides, if there is an elevated flux of 27-hydroxycholesterol into the brain as a consequence of a disturbed BBB, there may also be an increased flux of other oxysterols. Therefore, it may be assumed that in MCI or “compensated” AD stages, serum oxysterol levels are initially unchanged or slightly increased despite of presence of pro-oxidant and inflammatory stressors. However, in the “de-compensated” AD state, oxysterol homeostasis is overwhelmed by failure of cholesterol homeostatic mechanisms and widespread neuronal degeneration. These results highlight the importance of elaborating oxysterol profile changes among the different stages of AD so as to prevent or decrease neuronal damage.
The study taken here identified numerous relationships of serum levels of oxysterols with blood lipids, dietary cholesterol and fats and demographics. Common origins from cholesterol may explain many positive correlations with blood lipids. For example, the origin related to formation by enzymatic pathways may explain the consistent positive associations of 4β-hydroxycholesterol and 24-hydroxycholesterol with TC in both two batches. Besides, the consistent strong positive associations of 24-hydroxycholesterol with both HDL-C and LDL-C may also underlie the previously reported[41] cotransport of 24-hydroxycholesterol mainly with lipoprotein particles of HDL and LDL in the circulation. In addition to endogenous sources, oxysterols may also derive from the diet with food containing cholesterol and animal fat especially during food storing and cooking[42]. Findings in the validation batch including subjects with relatively higher dietary cholesterol and fats intakes demonstrated most positive correlations of oxysterols with dietary variables. Unexpectedly, consistent negative correlations of 4α-hydroxycholesterol and inverse correlations of 27-hydroxycholesterol with dietary cholesterol were observed, suggesting the dietary sources may be overwhelmed by endogenous sources. Taking into account that oxysterols can be further metabolized to bile acids, they play an important role of balance between cholesterol absorption and excretion[43].
A majority of positive correlations between all pairs of oxysterols were identified in the discovery batch, where smaller number of negative correlations were also presented. Shared metabolic pathways and precursor-product relationships may explain stronger positive correlations. However, the validation batch with increased MCI cases demonstrated more negative correlations between oxysterols. Consequently, we report emerging negatives correlations between oxysterols in MCI patients and the results suggested that the changes of interplay between oxysterols may be important indicators of cognitive status.
Genetic association analyses revealed significant associations between variants in genes encoding proteins or enzymes that are known to transport, synthesize and metabolize the oxysterols and cognition, blood lipids and oxysterol levels. Previous genetic association studies in AD or MCI have robustly identified several genetic risk variants in APOE and CYP46A1[44, 45], the latter of which specifies sterol 24-hydroxylase in the brain. In our analyses, five genetic variants in APOE, CYP46A1 and CYP7A1 were associated with decreased MCI risk while another three variants in CYP46A1 with increased MCI risk in specific genotype models, highlighting the heterogeneous effects of different loci in CYP46A1 for MCI development. We also investigated the association of blood lipids and serum oxysterol profiles with genetic variants in six oxysterol-related genes. On one hand, the inverse associations of rs6436087 in CYP27A1 with MMSE score and 7α-hydroxy-3-oxo-4-cholestenoic acid levels suggested that CYP27A1 may influence the risk of MCI through 27-hydroxycholesterol metabolism or changes in the brain. On the other hand, the consistent associations of rs3808607 in CYP7A1 with DSF score and 7α-hydroxy-4-cholesten-3-one indicated that CYP7A1 enzyme that catalyzes an essential step in bile acid synthesis[46] may influence MCI risk through cholesterol excretion. It is noteworthy that the seemingly protective role of genetic variants in APOE contrary to previous consensus[47], especially APOE4, may be due to significantly lower distribution of risk alleles of APOE ε4 in our population. These results should be interpreted with caution owing to ethnic homogeneity[48].
A limitation is the cross-sectional nature of the case-control studies, and therefore we can only detect correlations between clinical phenotypes and oxysterol levels but not draw causal conclusions. Longitudinal design would be better for in-depth examinations of dynamic changes of oxysterol biomarkers and predictive ability for the progression from prodromal to probable AD. Another limitation is lack of comparison with AD patients, thereby limiting the application of oxysterol biomarkers for discriminating among the different stages of AD, which may be more helpful to clarify dynamic changes of oxysterols and develop precise and targeted preventive and therapeutic strategies at the appropriate time.
{kind=link}