Identification and classification of SnRK2 genes in Rosaceae
To investigate the SnRK2 gene family in Rosaceae, the protein sequences of SnRK2s from Arabidopsis and rice were used as queries. Moreover, an Hidden Markov Model (HMM) search using the SnRK2 gene domain HMM profile (PF00069.1) was used to screen the Rosaceae genome, respectively. A total of 318 candidate SnRK2 genes were screened using these two strategies. Finally, the online program SMART (http://smart.embl-heidelberg.de/) was used to check the Ser/Thr protein kinase catalytic domains, followed by the removal of redundant sequences, incomplete gene sequences, and transcripts of the same gene. Subsequently, 71 nonredundant SnRK2 genes were identified in the Rosaceae genome. Among them, 10 SnRK2 proteins were identified in Chinese white pear (PbrSnRK2s), 14 in apple (MdSnRK2s), 8 in strawberry (FvSnRK2s), 7 in peach (PpeSnRK2s), 7 in Chinese plum (PmSnRK2s), 5 in black raspberry (RocSnRK2s), 7 in cherry (PavSnRK2s), and 13 in European pear (PcSnRK2s) (Fig. 1).
To classify and investigate the evolutionary relationships among SnRK2 genes, a phylogenetic tree was constructed using multiple sequence alignment of SnRK2 protein sequences from the eight Rosaceae species, Arabidopsis, and rice. The results of this analysis showed that the SnRK2 gene family was clustered into three well-supported clades (Groups I, II, and III; Fig. 1), which is consistent with the findings of a previous study performed in Arabidopsis and rice [15]. Among them, 22 members belonged to Group I, 25 to Group II, and 24 to Group III (Fig. 1).
Characteristics of the SnRK2 genes in Rosaceae
To study the functions of the SnRK2 proteins, we performed systematic analysis of the physicochemical properties of the SnRK2 proteins in Rosaceae. We found that the SnRK2 protein sequences ranged from 198 to 891 amino acids, and that most of them contained 220 to 402 amino acids. The isoelectric point of 87.3% of the SnRK2 proteins was acidic, which indicates that SnRK2 proteins from Rosaceae are rich in acidic amino acids. Moreover, the molecular weights of these proteins ranged from 30.09 to 85.3 kDa (Table 1). The negative and positive GRAVY scores of proteins reflect their hydrophobicity and hydrophilicity, respectively [24]. The grand average of the hydropathy scores of all SnRK2 proteins was negative in Rosaceae, which indicates that these proteins are hydrophilic. In addition, we found that the aliphatic index ranged from 80.76 to 94.08 for the SnRK2 proteins from Rosaceae, which indicated that all of them are thermally stable (Table 1).
Structural diversity of SnRK2 genes in pear
The analysis of the arrangement of introns and exons can provide unique insights into the evolution and functions of gene families [25]. To better understand the evolutionary model of PbrSnRK2 genes, exon/intron analyses were performed by aligning genomic sequences with their corresponding coding domains from SnRKs in pear and Arabidopsis. The number of exons identified in the members of the SnRK2 gene family ranged from 4 to 10 in pear and Arabidopsis (Fig. 2). Most members of the individual groups exhibited a different number of exons/introns and a varying length of the coding sequence in pear, which confirmed the phylogenetic classification of SnRK2 genes. For example, SnRK2 genes in subgroup I in pear contained 9 exons. Most members of subgroup II included 5–6 exons, except PbrSnRK2.4 containing 9 exons. In addition, subgroup III contained a number of exons (i.e., 7–8), except PbrSnRK2.10 containing 4 exons (Fig. 2). Thus, the conservation of the number of exons in each subgroup strongly supports the close evolutionary relationship of PbrSnRK2 genes. This may be the result of replication events in the evolution of the gene family, which means that these subgroups originated via different evolutionary paths.
Sequence alignment of PbrSnRK2 genes
To understand the functional characteristics of PbrSnRK2 genes and their conserved domains, multiple sequence alignment of PbrSnRK2 and AtSnRK2 proteins was performed. It was found that all members of the PbrSnRK2 family have two conserved kinase domains in the N-terminal regions: an ATP-binding signature containing a lysine residue as an ATP-binding site and a Ser/Thr protein kinase active site signature (Fig. 3). In addition, the C-terminus included two distinct domains, which were termed Domain I and Domain II. Domain I is necessary in all SnRK2s and is needed for activation by osmotic stress, whereas Domain II only exists in strongly ABA-responsive kinases [26].
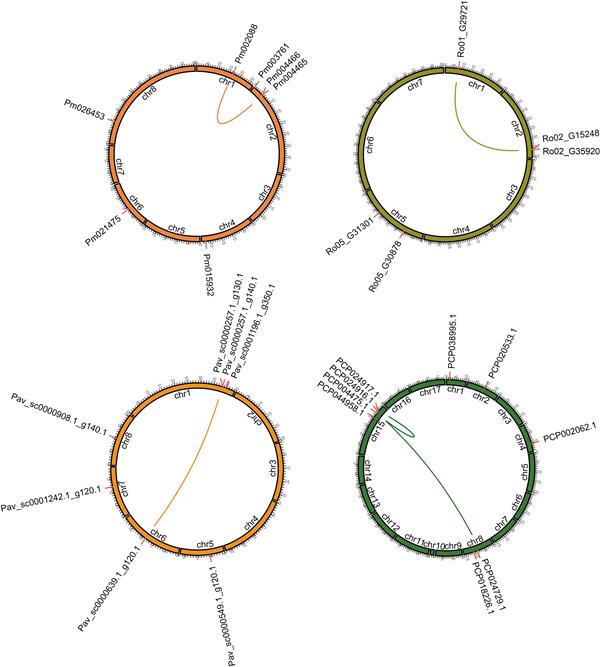
Synteny analysis of the SnRK2 genes in Rosaceae
To explore the evolutionary process of SnRK2 genes, a comparative syntenic map was constructed for Rosaceae. The PbrSnRK2 genes were distributed on 4 out of the 17 pear chromosomes, with 4 SnRK2 genes anchored on chromosome 15 (Fig. 4). Thirteen genes were assigned to 7 of the 17 chromosomes in apple, with 5 genes anchored to chromosome 15. In addition, 7 genes were distributed on 5 of the 8 chromosomes in peach, with 3 genes anchored to chromosome 1. Finally, 8 genes were assigned to 4 of the 7 chromosomes in strawberry, with 4 genes anchored to chromosome 5 (Fig. 4). Remarkably, similar to these observations, the SnRK2 genes in other Rosaceae species showed random chromosomal distributions (Fig. S1).
Analysis of the expansion of SnRK2 genes in Rosaceae
Several gene duplication patterns drive the evolution of protein-coding gene families, which include whole-genome duplication (WGD) or segmental duplication, tandem and segmental duplications, and rearrangements at the gene and chromosomal levels [27]. The origins of duplicated genes were explored in the SnRK2 gene family in eight Rosaceae genomes using the MCScanX package. Each member of the SnRK2 gene family was allocated to one of five different categories: WGD or segmental, singleton, proximal, tandem, or dispersed. Five types of duplication events contributed to the expansion of the SnRK2 gene family in Rosaceae: 50% WGD, 19.7% dispersed, 15.2% transposed, 9% proximal, and 6% tandem (Fig. 5). Among them, WGD events occurred in each of the Rosaceae species; in particular, 60% of the SnRK2 genes in Chinese white pear, 86% in apple, 57.1% in peach, and 50% in strawberry were duplicated and retained from WGD events compared with only 40% in black raspberry, 28.6% in Chinese plum, 28.6% in cherry, and 7.7% in European pear (Table S2). In addition, the proportions of dispersed SnRK2 gene duplication events in black raspberry (40%), European pear (30%), peach (28.6%), strawberry (28.6%), cherry (28.6%), and apple (7%) were assessed (Table S2). Therefore, gene losses, genome rearrangements, and RNA- and DNA-based transposed duplications may have occurred in these species. These results indicate that WGD and dispersed duplication play key roles in the expansion of the SnRK2 gene family in Rosaceae.
Ks value and Ka/Ks ratio reveal dates and driving forces of evolution
Purifying selection (negative selection) is the process via which disadvantageous mutations are removed, whereas Darwinian selection (positive selection) accumulates new advantageous mutations and spreads them throughout the population [27]. To identify the selection process that drove the evolution of the SnRK2 gene family, the Ka value and Ka/Ks ratio of its paralogs were examined in the eight Rosaceae species based on coding sequences. We found that all values were <1 in the studied Rosaceae species (Fig. 6A), implying that this family underwent a purifying selection pressure during its evolution in Rosaceae and that its evolution was very conservative.
The Ks value is extensively used to evaluate the evolutionary dates of WGD or segmental duplication events. To explore the evolutionary dates of the duplication events among the SnRK2 gene family members, Ks values were analyzed in the Rosaceae species. The results showed that the Ks values for the SnRK2 gene pairs ranged from 0.107 to 4.0487 in Rosaceae; moreover, the WGD gene pairs PbrSnRK2.1–PbrSnRK2.2 (Ks, ~0.1028), PbrSnRK2.7–PbrSnRK2.8 (Ks, ~0.1363), PCP022180.1–PCP002062.1 (Ks, ~0.182), MD01G1035000–MD15G1321000 (Ks, ~0.1644), MD02G1166500–MD15G1279000 (Ks, ~0.1757), MD04G1054400–MD06G1046300 (Ks, ~0.1635), MD08G1187200–MD15G1373000 (Ks, ~0.1138), and MD08G1236500–MD15G1428500 (Ks, ~0.107) indicated that some SnRK2 genes underwent WGD events more recently (30–45 MYA). Furthermore, other duplicated gene pairs (such as PbrSnRK2.5 and PbrSnRK2.6) possessed higher Ks values (2.26–4.0487), indicating that they might have stemmed from a more ancient duplication event (Fig. 6B).
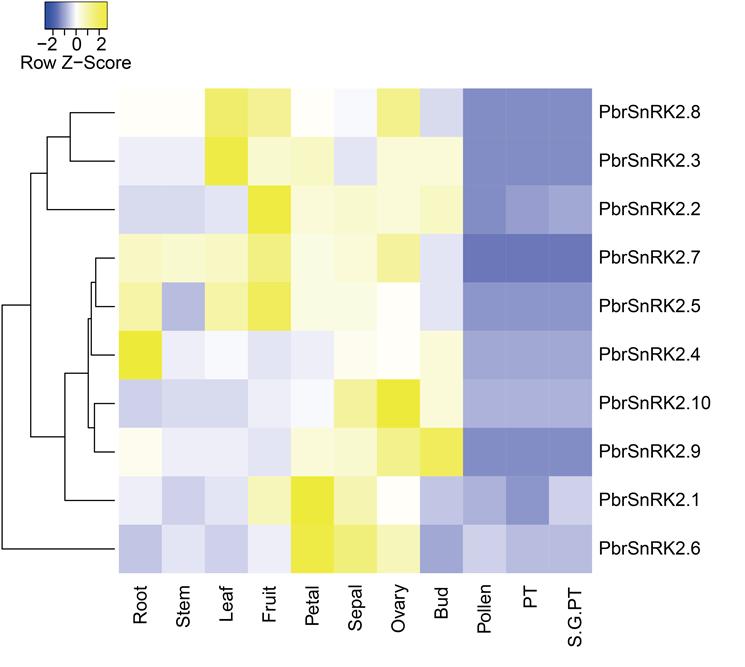
Expressions of PbrSnRK2 genes in different tissues
To understand the expression patterns and functional properties of SnRK2 genes in different tissues, we constructed a heat map at the transcriptional level using MeV to depict the overall expression patterns of SnRK2 genes in pear. The results showed that the SnRK2 genes were expressed in most organizations of pear, i.e., three genes (PbrSnRK2.2/2.5/2.7) were mainly expressed in fruits, two (PbrSnRK2.3/2.8) in leaves, and one (PbrSnRK2.4) in roots (Fig. S2). In addition, to validate the reliability of SnRK2 gene expression patterns based on the transcriptome data from pear, these SnRK2 genes were analyzed by qRT-PCR using gene-specific primers (Table S1). The results showed that SnRK2 genes were expressed in all tissues of pear, although there were differences in the absolute fold changes in the expression patterns between the two methods. For example, most genes were expressed in leaves and flowers, which indicated that SnRK2 genes mainly play important roles in these structures and respond to abiotic stress; in addition, two genes (PbrSnRK2.1/2.2) were mainly expressed in pollen and may play roles in regulating pollen germination (Fig. 7).
Expression profiles of PbrSnRK2 genes under ABA treatment
Many studies have shown that SnRK2s play key roles in response to multiple abiotic stresses such as salinity, dehydration, and hyperosmotic stress [14]. Moreover, the members of the SnRK2 gene family are involved in the regulation of phytohormone pathway responses, particularly ABA signal transduction [4]. To explore the dynamic transcriptional changes in PbrSnRK2 genes in response to ABA treatment, the expression levels of these genes in leaves of pear were evaluated by qRT-PCR under ABA (50 μM) treatment at four time points: 0, 3, 6, and 9 h. The results showed that in response to exogenous ABA application, the SnRK2 genes exhibited different expression patterns. For example, eight genes (PbrSnRK2.1/2.2/2.3/2.4/2.6/2.7/2.8/2.9) were activated by ABA, whereas the expressions of two genes (PbrSnRK2.5/10) remained unchanged at each time point (Fig. 8). Among them, the expression levels of PbrSnRK2.2/2.3/2.6/2.7/2.9 were significantly upregulated at 9 h after ABA treatment. PbrSnRK2.1 had a waving trend with ABA treatment, whereas PbrSnRK2.8 was downregulated over time after ABA treatment.
{kind=link}
{kind=link}