Domestication, breed formation, and selective breeding leave detectable patterns within the genome of livestock species such as sheep and goats. Identification of these genomic patterns in the DNA sequence could help to identify of genes controlling resistance to H. contortus or other gastrointestinal parasites. Several studies have attempted to identify the genetic variation controlling gastrointestinal parasite resistance in sheep and goats by using SNP markers and Genome Wide Association Studies (GWAS) but few research studies has been devoted to identify signatures of selection for GNI resistance in these species (27-29). Signatures of selection for resistance to GNI have not been identified in goats, and for sheep, only Perendale and Romney breeds have been evaluated (24). In the present study, we unravel signatures of selection using a targeted sequencing approach in three different breeds of sheep and goats. The SNPs potentially under selection identified in this study spanned a myriad of candidate genes related to immune response and cellular mechanisms against H. contortus.
In our study, all the signatures of selection identified in sheep and goats were under directional selection. Directional selection is one of the primary cause of phenotypic diversification and has been used to increase the frequency of favorable additive alleles (30). This selection process has not exhausted the genetic variation for most economically important traits in livestock (31). Our results suggest that some of the SNPs in genes related to resistance to GNI are under directional selection. This could be possible due to selection for resistance to GNI is focused on resistant individuals, and susceptible animals are usually removed from the flock or not used in the mating process.
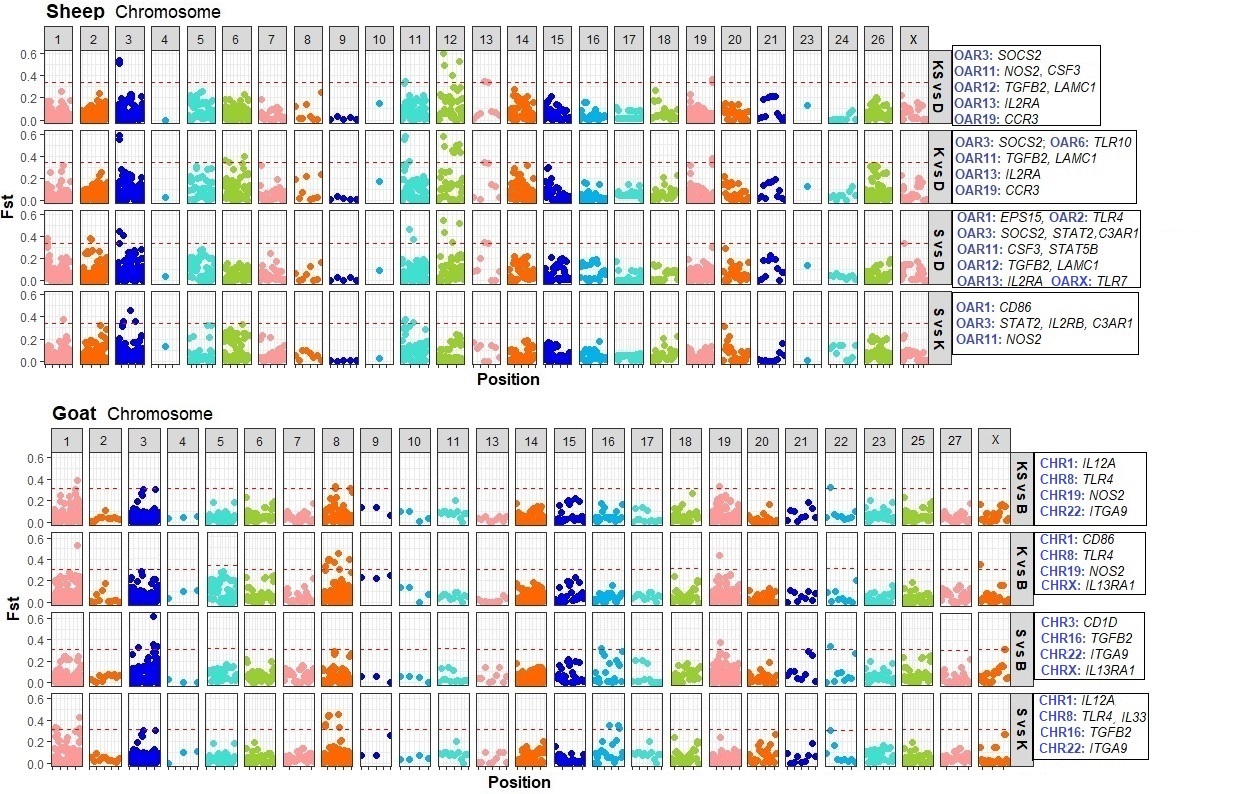
Signatures of selection in sheep populations
For Katahdin vs Dorper, and Katahdin and St. Croix vs Dorper analyses, loci within SOCS2, NOS2, TGFB2 and IL2RA genes were observed under selection. The SOCS2 gene has been previously associated with FEC in Dorper x Red Maasai sheep using GWAS and expression of this candidate gene was observed in abomasal tissue, mesenteric lymph nodes, and Peyer's patches from ewes and lambs (27). Thus, it is possible that SOCS2 gene could be used as candidate gene for future studies to validate previous and current results in Dorper and Dorper × Red Maasai sheep.
The SOCS2 gene is a broad key regulator of cytokine responses, including IL2, IL3, IL4, IFN-γ, CSF, and Jak-STAT signaling pathways in bone marrow and T cells (32). Studies on mice infected with Tripanozoma cruzi have shown that expression of SOCS2 facilitates inflammatory and immune responses to prevent myocardial dysfunction but increases parasitemia (33). On the contrary, SOCS2-/- mice infected with Schistosoma mansoni expressed increased Th2 response with higher IgE and eosinophil production than SOCS2+/+ mice (34). Also, SOCS2-/- mice have shown increased body weight and gigantism possibly due to downregulation of growth hormone and insulin-like growth factor-I (IGF-I) signaling (35). In Scottish Blackface sheep infected with Teladorsagia circumcincta, SOCS2 gene was found differentially expressed between resistant and control animals (36).
Nitrogen oxygen synthase 2 or NOS2 is a key molecule involved in Th1 response. It participates in the production of nitric oxide to kill invading microbes in phagocytes during classical macrophage activation by IFN-γ and TNF-α. Differential expression of this gene has been observed in the abomasum of Merino sheep during H. contortus challenge (37). In that study, mRNA expression NOS2 gene was downregulated in susceptible individuals. While there is a proposed interplay between Th1, Th2, and Treg responses during GNI (38), susceptibility to these infections has been related to Th1 and Th17 responses, and Th2 has been associated with resistance to helminth infections in sheep (39).
TGFB2 protein has been reported as an anti-inflammatory cytokine, and was observed in high concentration in the gut mucosa of sheep after H. contortus infection (40). In pigs, PAS1, a product of Ascaris suum, induces IL-10 and TGFB2 production in macrophages and has been related to loss of pro-inflammatory cytokine production (41). In humans and animal models, it has been shown that inhibition of T-cell proliferation might be triggered by an increase of IL-10 and TGFB production in antigen presenting cells or T-cells as a result of down-modulatory molecules that are released by the parasites to enhance survival (42). Thus, parasites are prone to use IL-10 and TGFB to downregulate host immune response.
IL2RA protein is mainly expressed in CD4+ Treg cells and it constitutes one of the three subunits of the IL2R. In humans, induction of Treg cells increases during natural and long term gastrointestinal nematode infections (43-44). In sheep, expression of the mRNA of IL2RA gene in the abomasum has been related to subsequent H. contortus infections in resistant sheep while its expression in the jejunal mucosa has been linked to susceptibility of Trichostrongylus columbriformis (37). Thus, differential expression between susceptible and resistant individuals could depend on the stage of the host immune response, the infection period, as well as the nematode species.
For St. Croix vs Dorper, and Katahdin and St. Croix vs Dorper analyses, SNPs in C3AR1, CSF3, STAT5B, TGFB2 and IL2RA genes were found to be under selection. C3AR1 protein plays an important role during innate immune responses. It is part of the complement cascade. Reduced T cell responses has been observed when host animals lack of C3AR (45). Recent work in mice, using bone marrow transplant and RNA Seq analysis, identified that signaling by C3AR mediates macrophage recruitment after induced injury with cardiotoxin and muscle regeneration (46). The exact role of C3AR1 has not been evaluated in sheep during gastrointestinal nematode infections, but some studies suggest that the complement activation is one of the first mechanisms of protection against helminth infections (47) and classical and alternative complement pathways can be activated in resistant sheep to H. contortus (48).
The STAT5 gene can be activated by many cytokines such as GM-CSF and thymic stromal lymphopoietin (TSLP) in the dendritic cells. Activation of STAT5 by TSLP has been shown to trigger Th2 responses at barrier surfaces (49). Also, STAT5 signaling has been related to many biological processes, such as TCR signaling and basal proliferation of naïve CD4+ T cells (50). Moreover, STAT5B mediates the signal transduction of IL2, IL4, CSF1, and different growth hormones. Thus, it is possible that STAT5B gene is responsible for many cellular functions during H. contortus exposure and further analysis is required to confirm our results.
Signatures of selection in goat populations
For many years, there has been a debate about the immune mechanisms controlling H. contortus infections in sheep and goats. The same helminth species can parasitize both species but previous studies suggest higher levels of infection in goats (51).
For Kiko and Spanish vs Boer, genes with genetic differentiation were IL12A, TLR4 and ITGA9. IL12 protein is a major cytokine that controls the maturation of CD4+ T cells into Th1 cells and promotes IFNG production in response to intracellular parasites. IL12 protein is composed of an alpha chain (p35 subunit) and a beta chain (p40 subunit) linked by a disulfide bond (52). In Nelore cattle, some studies have suggested that susceptibility to gastrointestinal parasites is associated with an increase of Th1 response with high elevated worm burden and elevated IFNG and IL12 production (53).
Toll-like receptors (TLR) are vital for the detection of invading pathogens and are commonly expressed in antigen presenting cells and other immune cells (54). In resistant sheep infected with H. contortus and T. colubriformis, upregulation of several TLR genes, including TLR4, was observed in the abomasum. In the same study, susceptible individuals presented lower expression of this gene (37). Contrary to sheep, susceptible Angus yearlings infected with Ostertagia, Cooperia and Nematodirus spp, TLR4 showed higher expression in the mesenteric lymph nodes (55). In goats, increased expression of TLR4 gene in blood has been observed during inclusion of Sericea lespedeza in the diet (56). This observation could be related to biologically active tannin fractions from plants containing tannins such as S. lespedeza. Several studies have shown plant tannins are able to modulate the innate immune response and act as γ⸹-T cell agonists (56-57)
The ITGA9 gene encodes an alpha integrin that compose the integral membrane glycoproteins that mediates cell-cell and cell matrix adhesion. In resistant Merino sheep infected with H. contortus, transcriptome analysis results revealed ITGA9 gene as part of an enriched gene set related to the extracellular matrix receptor interaction pathway (58). The exact role of ITGA9 gene in goats is unknown, but further analysis could help to understand possible mechanisms of protection against H. contortus and other gastrointestinal parasites.
For Kiko vs Boer, a SNP in CD86 gene was observed under selection. This gene encodes a membrane bound protein in antigen presenting cells that binds CD28 and CTLA-4 proteins localized in the T cell membrane. Binding with C28 leads to T cell proliferation and cytokine production, while binding with CTLA-4 negatively regulates T cell response (59). Thus, it is possible that CD86 controls some T cell mechanisms in goats.
For Spanish vs Boer analysis, signatures of selection were identified in CD1D, TGFB2, ITGA9 and IL13RA1 genes. CD1D is a major histocompatibility complex class I related protein that regulates presentation of glycolipids antigens to natural killer T cells (60). In resistant cattle naturally exposed to gastrointestinal parasites, CD1D was upregulated in the mesenteric lymph nodes (55). In goats, the role of CD1D has not been studied but it is possible that this gene could play an important role during presentation of glycoproteins from H. contortus to T cells.
As observed in sheep, TGFB2 could be used by H. contortus to promote infection. In tropical cattle, susceptibility to Theileria annulata has been associated with TGFB2 induction and increased TGF-b2 production by Theileria-infected macrophages promote invasiveness (61). In sheep, TGFB-like molecules have been identified in larvae from H. contortus and T. circumcincta (62). Thus, activation of TGFB and TGFB-like molecules from gastrointestinal parasites could control downregulation of the immune response. The exact role of TGFB2 in goats is unclear and more research is needed to understand its function during H. contortus infections.
IL13RA1 subunit together with IL4RA can form a functional receptor for IL13 (63). In goats, no evidence of IL13RA1 function has been reported but in Hereford Shorthorn cattle infected with Boophilus microplus, results showed that IL13RA1 precursor was differentially expressed (64).
Finally for Spanish vs Kiko, TLR4 and ITGA9 had SNPs under selection. For this analysis, the same SNP identified under selection in ITGA9 gene in Kiko and Spanish vs Boer analysis was observed. For TLR4, 2 more SNPs in exon 4 were identified under selection. Thus, it seems that for goats, TLR4 and ITGA9 genes could play important roles during H. contortus infection.
Common signatures of selection in sheep and goats
During the last two decades, results have shown differences in feeding behavior and gastrointestinal nematode parasitism between sheep and goats. Feeding behavior is one important aspect that differentiates sheep and goats. Sheep are typically raised in grazing systems with parasites commonly found within the pastures, and have to counteract the negative effects of GNI by developing an effective immune response. Goats are common browsers which allows them to rely less on immune response mechanisms (65).
For many years, there has been a question of the importance of immune effector molecules and the mucosal response in goats during GNI. Our findings suggest a possible interplay between Th1 and Th2 responses with conserved breed specific mechanisms. For both species, our results suggest a possible interplay between Th1 and Th2 response during GNI, as previously described by Hassan et al. (39) and Pernthaner et al. (66).
One of the most interesting findings from this study is the identification of shared immune response mechanisms between sheep and goats (Figure 3). It is possible that some immune response mechanisms are shared between both species and induce an effective immune response against H. contortus. The NOS2, TGFB2, and TLR4 genes, observed under selection in both species, are key modulators of Th1 and Th2 responses, and active players of antigen recognition. Several studies in cashmere and dairy goats have evaluated the responsiveness of resistant animals to GNI and have found a negative correlated response between worm counts and eosinophil, mast cell, and globule leucocyte counts (67-69). In sheep, similar cellular immune response has been associated to GNI (70-71). Thus, it is possible that some mechanism of immune protection are shared between these species but more studies are required to understand these events during H. contortus infection.
The conserved mechanisms of protective response against H. contortus are most likely to be useful targets in the development of alternative nematode control strategies in both species, as they can be widely applied in production systems. For this reason our future efforts will focus on validation of the results observed in the present study to unravel genetic mechanisms used for controlling H. contortus or other GNI between sheep and goats.
{kind=link}
{kind=link}