Recombinant RBD protein of SARS-CoV-2 exhibited good conformation and antigenicity. To obtain immunogen for immunization of mice, we constructed an expression vector encoding SARS-CoV-2 RBD and Fc fragment of human IgG (Figure 1a). After expression in Expi293F cells and purification with affinity chromatography, we characterized the RBD-Fc protein in both boiled and unboiled conditions using SDS-PAGE. As shown in Figure 1b, a single strong band with a molecular weight of ~60 kDa was noted in the boiled condition and one of ~120 kDa in the non-boiled condition, suggesting that the well-expressed RBD-Fc with high purity had formed a dimer via the Fc in the non-boiled condition. The recombinant RBD-Fc protein expressed in high yields, and the purification was high.
Using ELISA to investigate the binding activity between RBD-Fc and human ACE2 (hACE2), we found that the RBD-Fc could effectively bind to the hACE2 in a dose-dependent manner with a 50% effective dose (EC50) of 0.27 µM (Figure 1c). These results indicate that RBD-Fc exhibits proper conformation and functionality and that it can serve as an excellent immunogen.
RBD-Fc induced strong RBD-specific IgG antibody responses in mice. Three different doses of RBD-Fc (10, 5 and 2.5 µg, respectively) were used to immunize mice in order to optimize the dose of antigen that could sufficiently induce antibody response. The mice were vaccinated subcutaneously three times at 14-day intervals, and the sera were collected at 7 days after each boost (Figure 2a). The geometric mean titer (GMT) of RBD-specific IgG in sera was detected with ELISA using the protein of RBD-His, rather than RBD-Fc, to coat the plates in order to eliminate the influence of antibodies against Fc. For the sera collected from mice on day 21 post-1st immunization with 2.5, 5, and 10 µg of RBD-Fc, the GMTs of SARS-CoV-2 RBD-specific antibody IgG are 32,254, 64,508 and 117,713, respectively (Figure 2b and 2c), and those of SARS-CoV- 2 S1-specific antibody IgG are 3,200, 10,159 and 19,577, respectively (Figure 2d and 2e), while the sera from mice treated with PBS exhibited only background level of the signal. For the sera collected on day 35 post-1st immunization with 2.5, 5, and 10 µg of RBD-Fc, the GMTs of RBD-specific antibody IgG were increased to 315,420, 378,800 and 546,323, respectively, while those of SARS-CoV-2 S1-specific antibody IgG were increased to 126,267, 262,645, and 454,914, respectively (Figure 2f-2i). These results suggest that immunization of mice 2 to 3 times with RBD-Fc at 2.5 - 10 µg could elicit similarly strong RBD-specific antibody responses.
RBD-Fc elicited robust neutralizing antibody responses in mice against pseudotyped and live SARS-CoV-2 infection. To assess whether the sera contains neutralizing antibodies against SARS-CoV-2 infection, we developed a pseudovirus (PsV) expressing S protein of SARS-CoV-2, which could simulate the SARS-CoV-2 virion to enter and infect the target cell without replication28-30. The sera collected from mice immunized with 2.5, 5 and 10 µg of RBD-Fc at day 21 post-1st immunization potently neutralized SARS-CoV-2 PsV infection in Huh-7 cells with geometric mean 50% neutralization titers (NT50) of 864, 1,389, and 1,795, respectively (Figure 3a). The geometric mean NT50s of the sera were increased to 1,965, 8,179, and 7,166, respectively, at day 35 post-1st immunization (Figure 3b). These results suggest that immunization of mice with 5 µg RBD-Fc is able to elicit highly potent neutralizing antibody responses.
We then developed a competition ELISA assay to detect whether SARS-CoV-2 RBD-specific antisera could block the interaction between SARS-CoV-2 RBD and hACE2. As shown in Figure 3c, antibodies in the antisera from mice immunized with RBD-Fc at day 35 post-1st immunization could significantly inhibit the binding of SARS-CoV-2 RBD to hACE2, compared with the sera from mice treated with PBS, indicating that RBD-Fc-induced antibodies could neutralize SARS-CoV-2 PsV infection by blocking the binding of RBD in S protein on the surface of SARS-CoV-2 to hACE2.
Next, we tested the neutralization activity of the antisera collected from mice immunized with 10 µg RBD-Fc at day 35 post-1st immunization against live SARS-CoV-2 infection in Vero-E6 cells using an immunofluorescence assay31. After the addition of the antisera at a dilution of 1:2,700 or 1:8,100, we found that the expression of SARS-CoV-2 N protein in the cells was almost completely, or partially, suppressed, respectively (Figure 3d). Then, we further tested the viral RNA in the cell supernatant using an RT-qPCR assay. The results showed that the antisera could inhibit live SARS- CoV-2 infection in a dose-dependent manner with an NT50 of 10,523 (Figure 3e), consistent with the result from the immunofluorescence assay.
Recently, hACE2 transgenic (hACE2-Tg) mice were used for the development of an animal model of SARS-CoV-2 infection32. We, therefore, immunized hACE2-Tg mice with the same immunization strategy to investigate the protective effects of vaccination in vivo (Figure 2f). The hACE2-Tg mice were vaccinated with 5 µg RBD- Fc three times and then challenged with SARS-CoV-2 intranasally. The lungs were collected at 4 days post-challenge to test their virus titer. As shown in Figure 2g, the viral RNA was not detected in the lungs from RBD-Fc-immunized mice, while relatively high viral RNA copies were detected in the lungs from PBS-treated mice.
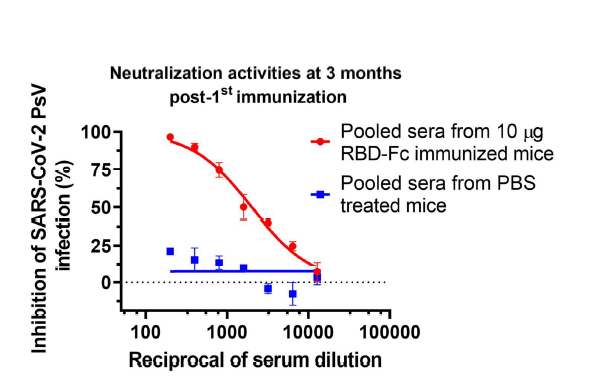
To investigate whether the neutralizing antibody response in mice immunized with RBD-Fc could maintain at high level for a longer period of time, we tested the neutralization activity of pooled sera from mice immunized with 10 µg RBD-Fc at 3 months post-1st immunization. The results showed that pooled sera could also effectively neutralize the SARS-CoV-2 PsV infection with the NT50 of 2,013 (Figure S1). These results indicate that the RBD-Fc vaccine is able to induce relative long-term neutralizing antibody responses.
RBD-Fc-induced antibodies inhibited SARS-CoV-2 S-mediated cell-cell fusion. Subsequently, we adapted a cell-cell fusion assay by utilizing 293T effector cells expressing SARS-CoV-2 S protein and EGFP and Huh-7 target cells expressing human ACE228, 29 to test whether RBD-specific antisera could block SARS-CoV-2 S protein- mediated cell-cell fusion. We found that all antisera from mice vaccinated with 2.5, 5 or 10 µg at a dilution of 1:100 could completely inhibit SARS-CoV-2 S-mediated cell- cell fusion, while the sera from the PBS-treated mice showed no significant inhibitory effect (Figure 4a). The calculated geometric mean IT50 (50% inhibition titers) of the sera collected from the mice vaccinated with 2.5, 5 and 10 µg at day 35 post-1st immunization were 724 (95% CI, 404 to 1294), 1,787 (95% CI, 734 to 4,348) and 1,503 (95% CI, 555 to 4071), respectively. Consistent with the results from SARS-CoV-2 PsV inhibition assay, the 5 µg group showed inhibitory activity against cell-cell fusion similar to that of the 10 µg group, but much better than that from the 2.5 µg group, suggesting, again, that 5 µg is the optimal dose of RBD-Fc for immunization. Collectively, these results show that immunization of mice with 5 µg of RBD-Fc can elicit high titers of RBD-specific antibodies to neutralize infection of pseudotyped and live SARS-CoV-2 and inhibit SARS-CoV-2 S-protein-mediated membrane fusion.
SARS-CoV-2 RBD-Fc could induce cross-neutralization of antibodies against infection of pseudotyped SARS-CoV-2 mutants, SARS-CoV and SARSr-CoV. SARS-CoV-2 has been undergoing continuous mutations in its S protein, especially in the RBD region33, which may render the virus resistant to neutralizing antibodies induced by the COVID-19 vaccines currently under development. To determine whether the mutations in RBD are resistant to the neutralizing antibodies induced by RBD-Fc, we constructed 7 SARS-CoV-2 PsVs with single natural mutation in RBD of SARS-CoV-2 S protein and used them to analyze their sensitivity to the neutralization activity of mouse antisera. All 7 mutants could be effectively neutralized by the pooled sera collected from mice immunized with 10 µg RBD-Fc at day 35 post-1st immunization with NT50 of >3,000 (Figure 5a-5h).
Given that the RBD of SARS-CoV-2 has 74.7% and 75.2% amino acid sequence identity to the RBD of SARS-CoV and bat-SARSr-CoV-WIV1, respectively (Figure 6a), we determined whether SARS-CoV-2 RBD-Fc-induced antibodies could cross- react with RBD of SARS-CoV using ELISA. As shown in Figures 6b and 6c, the sera collected from mice immunized with 10 µg RBD-Fc at day 35 post-1st immunization could effectively bind to RBD of SARS-CoV in a dose-dependent manner with IgG GMT of 51,200. We then purified the total IgG from the pooled sera collected from mice immunized with 10 µg RBD-Fc at day 35 post-1st immunization to test its neutralization activity against SARS-CoV-2 PsV infection. We found that total SARS- CoV-2 IgG could significantly neutralize infection of SARS-CoV-2 PsV (Figure 6d), SARS-CoV PsV (Figure 6e), and bat-SARSr-CoV PsV (Figure 6f), respectively, although the neutralizing activity against SARS-CoV-2 PsV turned out to be more potent than that against SARS-CoV PsV and bat-SARSr-CoV PsV. These results serve as additional evidence that SARS-CoV-2 RBD-Fc has the potential to be developed as a broad-spectrum vaccine to prevent infection by lineage B betacoronaviruses, including SARS-CoV-2 that has caused the current COVID-19 pandemic, as well as SARS-CoV and bat-SARSr-CoV that may cause the future coronavirus disease outbreaks.
Identification of the linear epitopes in the SARS-CoV-2 RBD domain. To investigate whether RBD-Fc-vaccinated mice could produce antibodies targeting linear epitopes in the SARS-CoV-2 RBD domain, we synthesized a group of peptides spanning the RBD region in S protein (residues 314-543). Each peptide contained 20 amino acids with 10 residues overlapping with the adjacent peptides. These peptides were precoated in a chip. The results revealed that four linear peptides in the SARS- CoV-2 RBD domain could react with the antisera (Figure 7a). To further confirm the reactions and quantify the binding capacity, we coated the four peptides on ELISA plates to detect antibody-peptide binding in sera. Some sera induced by RBD-Fc showed higher binding capacity to the four linear peptides (Figure 7b-7e). Particularly, peptide S464-483 could strongly react with the antisera collected from mice vaccinated with 2.5, 5 or 10 µg at day 35 post-1st immunization, demonstrating that S464-483 is an immunodominant linear epitope in the SARS-CoV-2 RBD domain.
We then tested whether these peptides could block the neutralizing activity of antibodies in mouse antisera. The results showed that the neutralization was not significantly affected by these peptides at the concentration as high as 10 µg/ml, while RBD-Fc at 0.078 - 0.312 μg/ml could effectively block the neutralization activity of RBD-specific antibody (Figure 7f). We then tested whether these peptides could inhibit SARS-CoV-2 PsV infection and found that none of these peptides could significantly inhibit SARS-CoV-2 PsV infection, whereas the RBD-Fc protein could effectively inhibit SARS-CoV-2 PsV infection in a dose-dependent manner (Figure 7g). These results suggest that most SARS-CoV-2 RBD-specific neutralizing antibodies recognize conformational, but not linear, epitopes. Some peptides containing linear epitopes may induce non-neutralizing antibodies, consistent with the finding of SARS-CoV RBD- specific neutralizing antibodies34. When designing a COVID-19 vaccine, the epitope with the ability to induce non-neutralizing antibody or neutralizing antibody with antibody-dependent enhancement (ADE) effect should be masked with a glycan probe as previously described35.
{kind=link}