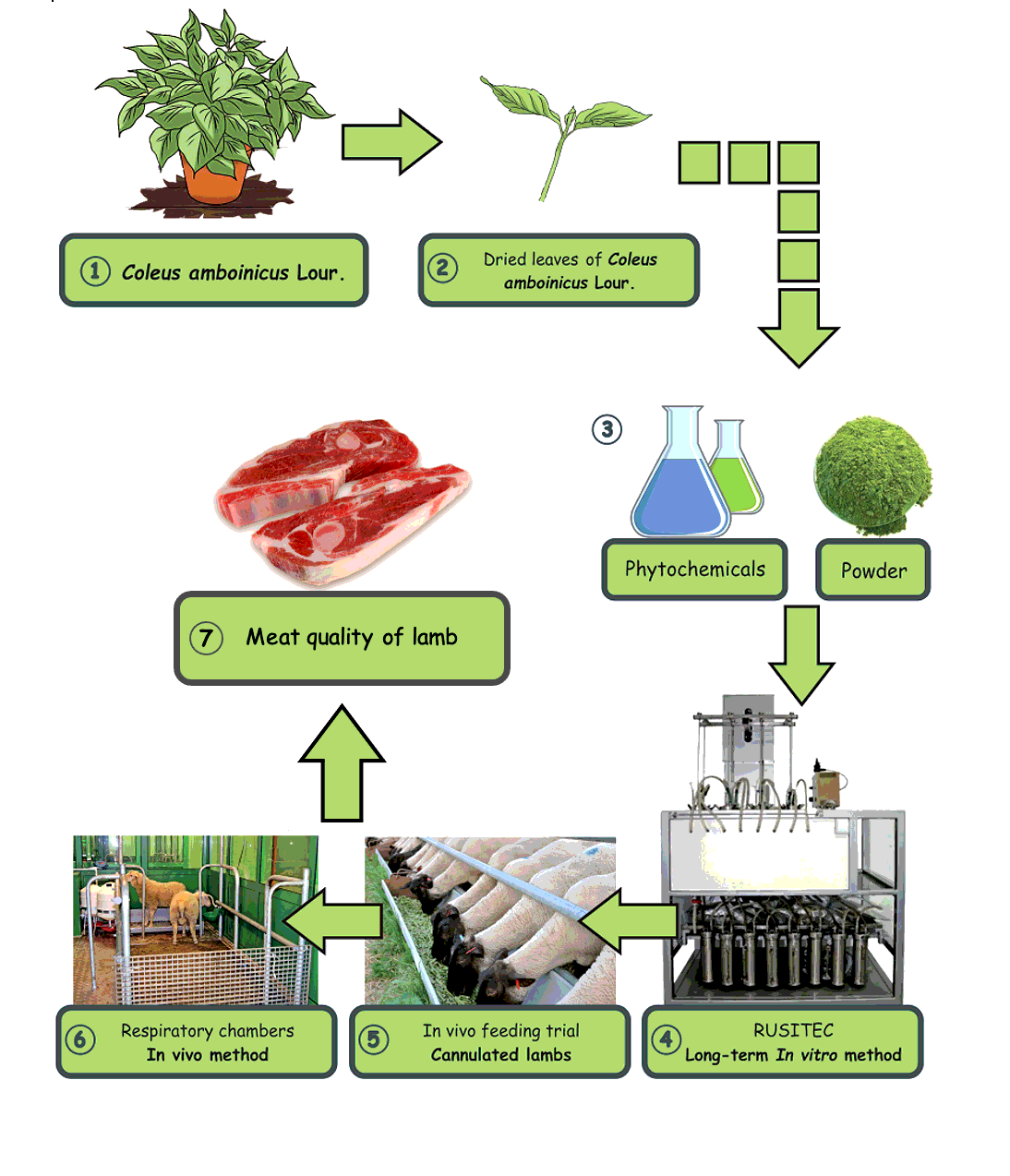
In vitro experiment (10-day RUSITEC fermentation, Experiment 1)
Identification of the concentration of particular BAC in the CAL undoubtedly helped to interpret our results. The gradual replacement of a proportion of the main diet (from 10–20%) by CAL decreased methane production and gradually diminished population of ruminal methanogens (both total methanogens and methanogen species observed). The greatest reduction in total methanogen population (by 24%) as well as the most effective mitigation of CH4 production (by 49%) was observed for the 20% CAL diet. As in our previous study [13], no negative changes in rumen parameters were noted. Our findings are also supported by other authors[41, 42], who demonstrated CH4 reduction in response to BAC of plant origin, with no negative consequences such as declined feed degradability, changes in ammonia, total VFA concentration, or microbial population (total protozoa and bacteria population). We have also noted the inhibition of methanogens via a direct toxic antibacterial effect of BAC, but without adverse effects on ruminal fermentation or DM degradability. Despite the toxic action of BAC on methanogens, no effect on protozoa and ruminal bacteria was observed. Only the lowest level of CAL (10%) increased the population of the bacteria Prevotellaspp., B. proteoclasticus, and B. fibrisolvens. A distinct effect of the lowest CAL supplementation was also observed in our previous study on Saponaria officinalis[43]. We hypothesized that the basic nutrient components of a plant can either interact with BAC or become physically less available for microbiota, resulting in a decreased antibacterial activity.Moreover, the reduction in the methanogen population resulting from the antimethanogenic effect of CAL lowered the acetate:propionate ratio, probably by shifting the free-hydrogen pathways to propionate production. Our findings corroborated with an experiment on Rosmarinus officinaliscontaining diterpenes (such as carnosol, carnosic acid, rosmanol, epirosmanol, isorosmanol, and methyl carnosate) added to a ruminant diet [44]. This supplement affected several groups of rumen microbes involved in protein and fiber degradation as well as CH4 and ammonia production. Coleus amboinicus (Lour.) is a rich source of UFA with the predominance of α-linolenic acid (LNA). The higher fat concentration in CAL than in grass silage (43 vs 21 g/kg DM, respectively) may increase the ruminal FA content, and consequently that of LNA. The effect of CAL in the diet on FA profile in the rumen differed from that on bacteria population. For the majority of FA, the significant effect was observed for all CAL (10%, 15%, and 20%) diets.
Dietary CAL modulated the ruminal FA composition by lowering SFA and elevating MUFA and PUFA proportions. The alterations in MUFA and PUFA profiles may suggest the changes in BH process in the rumen. We showed in our short-term study that the polyphenol compounds of CAL origin (flavonoid, phenolic acids, and diterpenes) can alter the action of the ruminal microbiota involved in BH [13]. We also noted higher content of total polyphenols and diterpenes of CAL origin in this long-term study compared with our previous short-term study [13]. The CAL diet decreased C18:1 trans concentration due to the BH process of LA. High quantities of polyphenols and diterpenes reduced the final BH step of the C18:1 cis-9 and caused a linear decrease in the concentration of stearic acid (C18:0). This was also observed by Vasta et al. [5], who described a negative effect of different polyphenols on C18:0 accumulation in the rumen digesta. The results of the current study thus demonstrate the protective action of CAL on BH of MUFA and PUFA.
In vivo experiments: Experiment 2 with cannulated lambs and Experiment 3 with growing lambs.
The many ruminal fermentation characteristicsin the growing lambs receiving 200 g/day of CAL corroborated with the results of the in vitro study. Methane emission (L/d and L/DM intake) was mitigated by 20% and 29%, respectively, and did not interfere with DM, OM, or NDF digestibility. Decreases in CH4 production are usually associated with adverse effects on fiber digestibility [45]. We suggest that, in this case, CH4 mitigation was attributed directly to the reduction in total methanogen and Methanobacterialespopulation (in both growing and cannulated lambs), as well asin Methanomicrobiales populations (in the cannulated lambs) rather than reduction of carbohydrate digestibility [41]. These results are supported by our previous short-term in vitro studies investigating BAC from two sources (Sanguisorba officinalis and CAL), where there were no negative effects on in vitro parameters, including DM degradability [13].
The utilization of forages in ruminants requires the coordinatedactivity of bacteria (such as F. succinogenes, R. albus, R. flavefaciens, and B. fibrisolvens), protozoa, and fungi for proper digestion[5, 46, 47]. Digestibility is also affected by the number and composition of rumen microorganisms. Supplementing the diet with 200 g/day of CAL had a positive effect on the total protozoa population in growing lambs (Experiment 3). The effect was more pronounced for a few particular bacterial species (namely R. albus, M. elsdenii, B. proteoclasticus, and B. fibrisolvens) than for the total bacteria. In comparison with tannins, BAC of CAL origin exerted either less negative impact on the rumen environment, or else their concentration was more beneficial on the ruminal microorganisms. This is supported by De Nardi et al. [48], who reported an increase in the number of several ruminal bacteria in heifers due to supplementation of flavonoids and essential oils.
Neither digestibility nor total VFA concentration was affected in the present study, although alterations in individual VFAs, such as propionic acid, were noted. The high proportion of propionate was not linked with the abundance of S. bovis, the amylolytic bacteria responsible for production of this VFA. The possibility cannot be however excluded that other amylolytic bacteria, not investigated in this study, were not affected. Additionally, higher concentration of propionic acid and lower acetic to propionic ratio can reflect in decreased hydrogen accessible to methanogens[49]. Considering the direct reduction in methanogen count caused by BAC of CAL origin, such a pathway has rather limited importance for mitigation of methanogenesis in this study.
Another aspect of this experiment that is worthy of attention is the changes observed in nitrogen metabolizing bacteria population (Prevotella spp., B. proteoclasticus, and B. fibrisolvens). CAL is a feed rich in total protein (214 g/kg DM), and is thus the perfect substrate for N metabolism in the rumen. We observed an increase in ruminal ammonia in response to CAL supplementation. It is known that tannins form complexes with proteins, reducing the ammonia level in the rumen fluid. In the present study, protein degradation was not likely limited by CAL. Besides, the higher protein content may influence the ammonia level.
The in vivo experiments had higher ruminal pH values. Our observation corroborated the result ofBalcells et al. [50], who observedhigher pH after supplementing dairy cow diet with flavonoids.The extent of rumen fermentation reflected by the total VFA suggests that CAL supplementation did not affect this variable (Experiment 2 and Exp.3). Microbial activity is affected by the properties of the substrate, and higher starch levels in the diet usually reduce the pH of the rumen fluid [51, 52]. The diets in the present study were rich in concentrate (400 g/day) and we expected a reduction in pH. According to Balcells et al. [50], flavonoid supplementation may improve rumen fermentation and reduce the incidence of rumen acidosis. Such activity of some flavonoids may be partially explained by the increased number of lactate-consuming microorganisms (e.g., M. elsdenii). Our studies also confirmed this activity of CAL flavonoids by demonstrating increased pH values in the lamb rumen. It should be realized that the CAL contained three types of BAC with distinct activities: flavonoids, diterpenes and phenolic acidsbut the mechanism described above is typical for flavonoids. Another mechanism that may stabilize the pH is the fluctuations of protozoa community. According to Hartinger et al. [53], protozoa incorporate starch granules that are not metabolized to organic acids quickly. Thus, protozoa protect the rumen from large drops in pH, and support stable fermentation conditions. The effect of CAL on rumen protozoa observed in the present study was ambiguous. The Entodiniomorphapopulation significantly increased (Table 11), whereas that of Holotricha diminished consistently (Tables 9, 11). Similar trends were described in our short-term study [13], so we can assume that this mode of CAL action is stable for a longer period.
The FA proportion of the rumen fluid was another parameter affected by supplementation with CAL. We noted a positive alteration in the ruminal PUFA proportion in the growing lambs, which can mainly be attributed to the increase in C18:3 cis-9 cis-12 cis-15 FAs in the rumen fluid and meat (Table 12). The CAL polyphenols likely increased the bypass of UFA. Although the majority of bacterial communities involved in BH increased in number, BH of C18 UFA into stearic acid (C18:0) was lower due to CAL supplementation. A similar result was reported by Vasta et al. [5], who, regardless of the type of polyphenol supplementation, observed a reduction in ruminal C18:0 and an increase in C18:1 trans-11 (VA) and C18:2 cis-9 trans-11 (CLA) isomers. Vasta et al.[5] also pointed out a range of actions of polyphenols on bacteria involved in the BH process. The stability of the main BH isomers and the distinct responses of two bacteria species involved in BH process (B. fibrisolvens and B.proteoclasticus) observed in our study can also demonstrate some other properties of polyphenols. The reduction of BH by CAL despite greater populations of BH bacteria may suggest that CAL may reduce the content of free FA, substrates to BH process due to decreased lipolysis [4]. Besides, a reduction in the numbers of several bacteria species involved in BH, such as Neisseria weaverii, R. amylophilus, and other unclassified bacteria related to the Lachnospiraceaeand Pasteurellaceaefamilies was observed [54]. In this study, the potential restriction on BH may be supported by the fact that the rumen and meat displayed similar alterations in the FA profile, which likely suggests that the rumen was the locus of the main changes.
Transcript analysis included a panel of five genes of known function in FA metabolism in ruminant muscles [55]. Three of the genes (FASN, SCD, and ELOVL5) control the de novo synthesis and elongation of FA,whereas the LPL and FADS1 genesare involved in FA transport. However, that considering the complexity of the entire gene expression process and the great variation seen in transcript lifespans, any conclusions must be drawn with caution. Meat from lambs fed the CAL diet was characterized by a significant reduction in the mRNA content of four genes (FASN, SCD, LPL, and FADS1),whereas no changes were observed for the ELOVL5 transcript. Alterations in the transcript expression of genes regulating lipid metabolism was not reflected in the profile of FA controlled by those genes. For example, the reduced mRNA level of the SCD gene was not accompanied by a lower level of C18:1 cis-9. According to Garnsworthy et al. [56], the level of C18:1 cis-9 in the fat of ruminant products is highly dependent upon the SCD gene controllingde novo FA synthesis. Besides, the lack of difference in C18:1 cis-9 concentration in response to CAL supplementation may suggest that the synthesis of endogenous FA was unaffected. On the other hand, the reduced transcript levels of the LPL gene may be associated with the decreased biosynthesis of MUFA. The lack of alteration in the mRNA content of the ELOVL5 gene regulating FA elongation and the increased n-3 concentration (mainly LNA) in meat may reflect n-3 metabolism in the rumen, rather than that in muscles. Considering the reduced transcript content of another two genes—FADS1 and FASN—this may be linked to the inhibition of the initial stages of the BH process.Pewan et al. [57]suggested that the FASN protein complex controls de novo biosynthesis of long-chain FA and affects FA deposition in meat, adipose tissue, and milk. The published evidence on the correlations between n-3 PUFA profile, the activity of lipogenic genes (such as FASN), and meat quality, however, is very limited [57].Nevertheless, the n-3 PUFA profile of the meat of lambs fed CAL was improved, which suggests that changes in the FA profile had already occurred in the rumen, leading to more PUFA being available to the tissue. Higher n-3 PUFA levels in meat are beneficial to human health and support cardiovascular, retinal, and brain functions [58, 59]. Positive changes in the FA profile of meat from the experimental lambs increased its quality. However, higher content of PUFAs could decrease the meat shelf life due to a rapid oxidization of FAs ensuing from two or more double bonds in their structure [60].
The oxidative process can shorten the shelf life of fresh meat and negatively affect its consumption by the formation of off-flavors and discoloration [61]. The sensor attributes, likes juiciness and taste, are generally associated with the consumers’ preferences[62]. In this study, we observed improved meat characteristics such as juiciness or taste. The better meat sensory quality combined with improved indices of thrombogenicity (TI) and atherogenicity (AI) indexes may help to encourage consumers to select quality meat in the future. Tannin-containing diets sometimes reduce [63] or unchanged [64] tenderness and juiciness of meat depending upon the doses. In this study, juiciness and flavor of lamb meat increased, which would be advantageous from consumer perspectives. It has been suggested that phenolic compounds may increase the activity calpains and accelerating the degradation of myofibrillar proteins during the postmortem conditioning of carcass, leading to increased juiciness and tenderness of meat [65, 66]. A study with Andrographis paniculata, an annual herb rich in polyphenols,reported that this herb increased the juiciness and tenderness of the LT muscle of goats [66]. Previous study reported that dietary polyphenols from Mimosa condensed tannins had no influence on meat lightness and colors of lambs, however the meat metmyoglobin was reduced [67]. Polyphenol inclusion in ruminants’ diet might cause different effects on the meat color. Many studies suggested that dietary polyphenols might cause a lighter color of meat [68, 69]. Priolo et al. [70] evaluated the effect of tannin in lambs and stated that the longissimus muscle was lighter (lower L*) in tannin-fed group. We assume that the lower lightness and greater redness of color in the present study were influenced by the CAL mineral content, which was not determined in this study. Damanik et al. [12] reported that the CAL contains high concentrationof iron (Fe). Garg et al. [71] stated that the inclusion of tannin in the animal diets do not hamper the utilization of iron for the hemoglobin synthesis. Thus, lambs fed CAL diet might influence its carcass color and lightness due to the high level of Fe affecting the hemoglobin synthesis in the meat. Moreover, polyphenols with high antioxidant activities may reduce the oxidation of myoglobin leading to increased the redness of meat [61]. Increased redness of meat has also been reported due to feeding of polyphenol-rich plants [66, 72].
{kind=link}