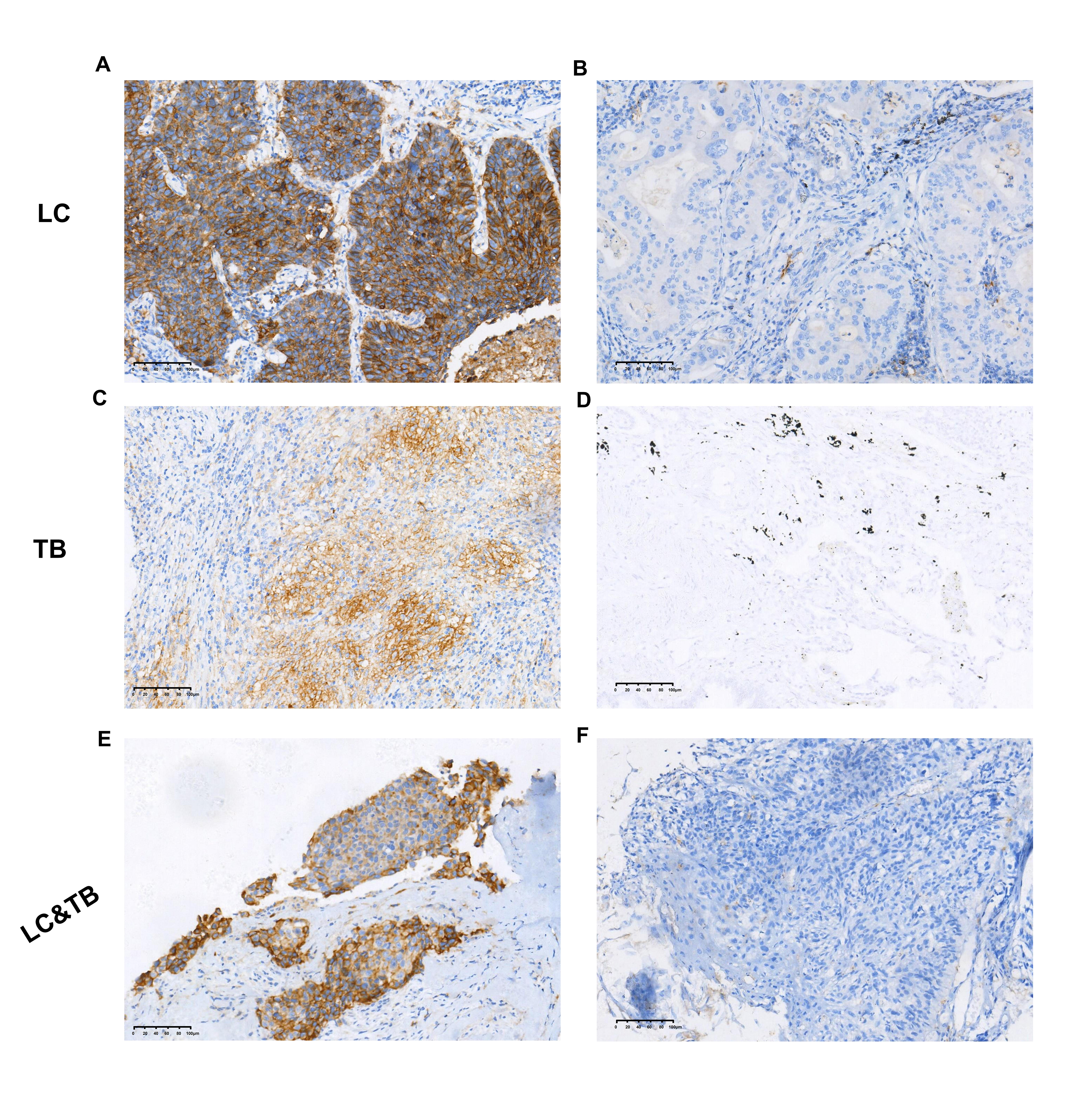
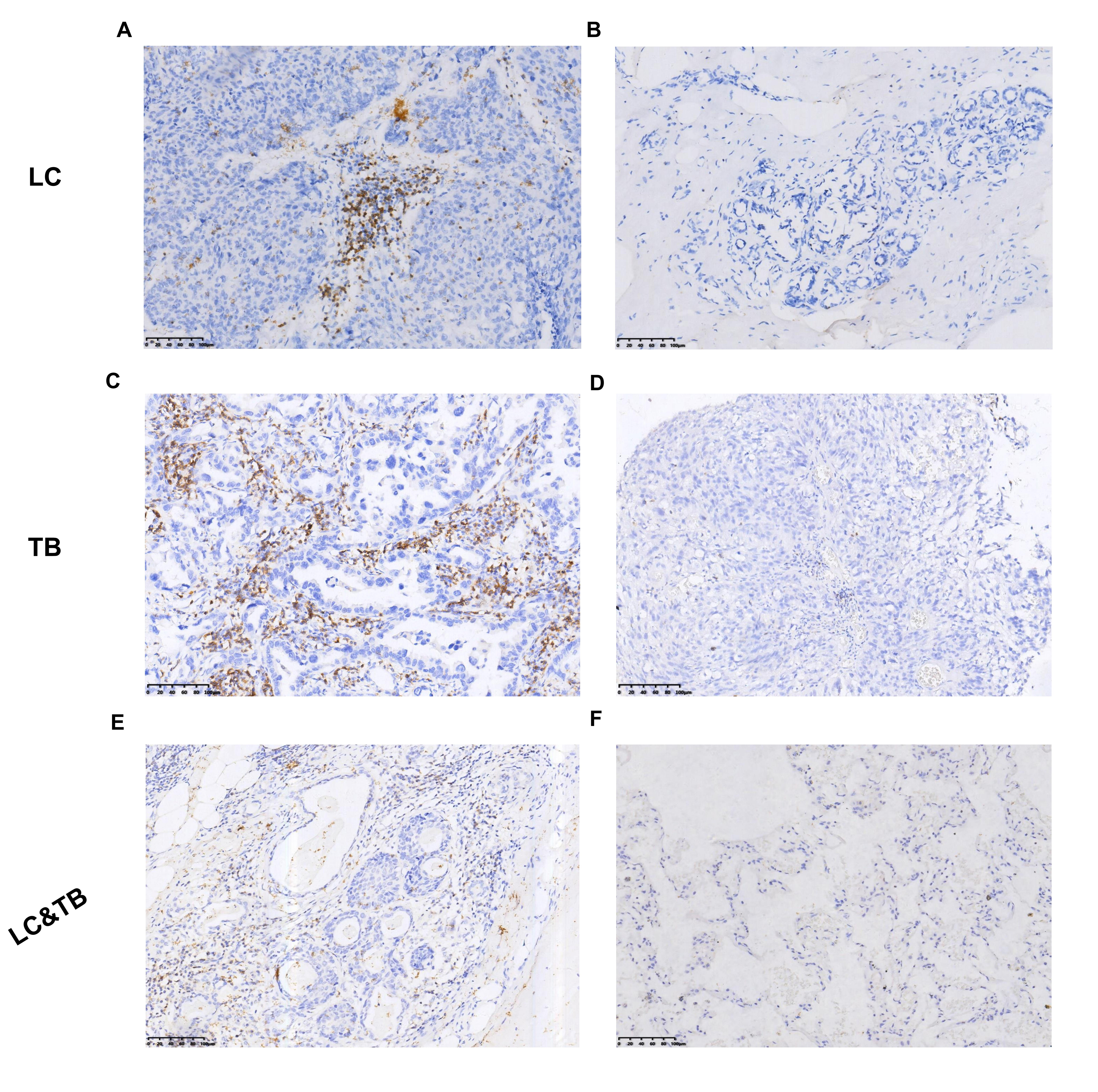
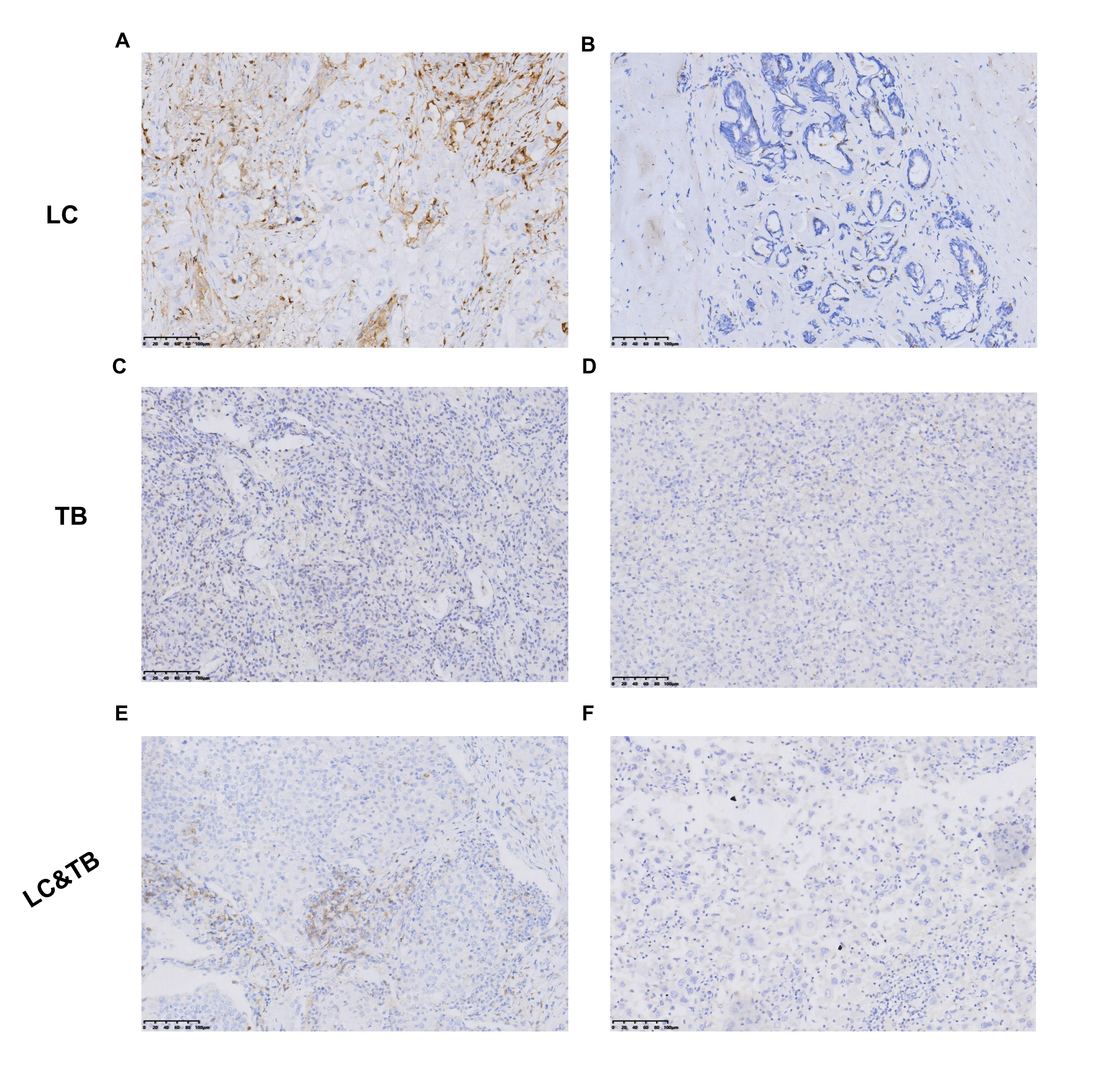
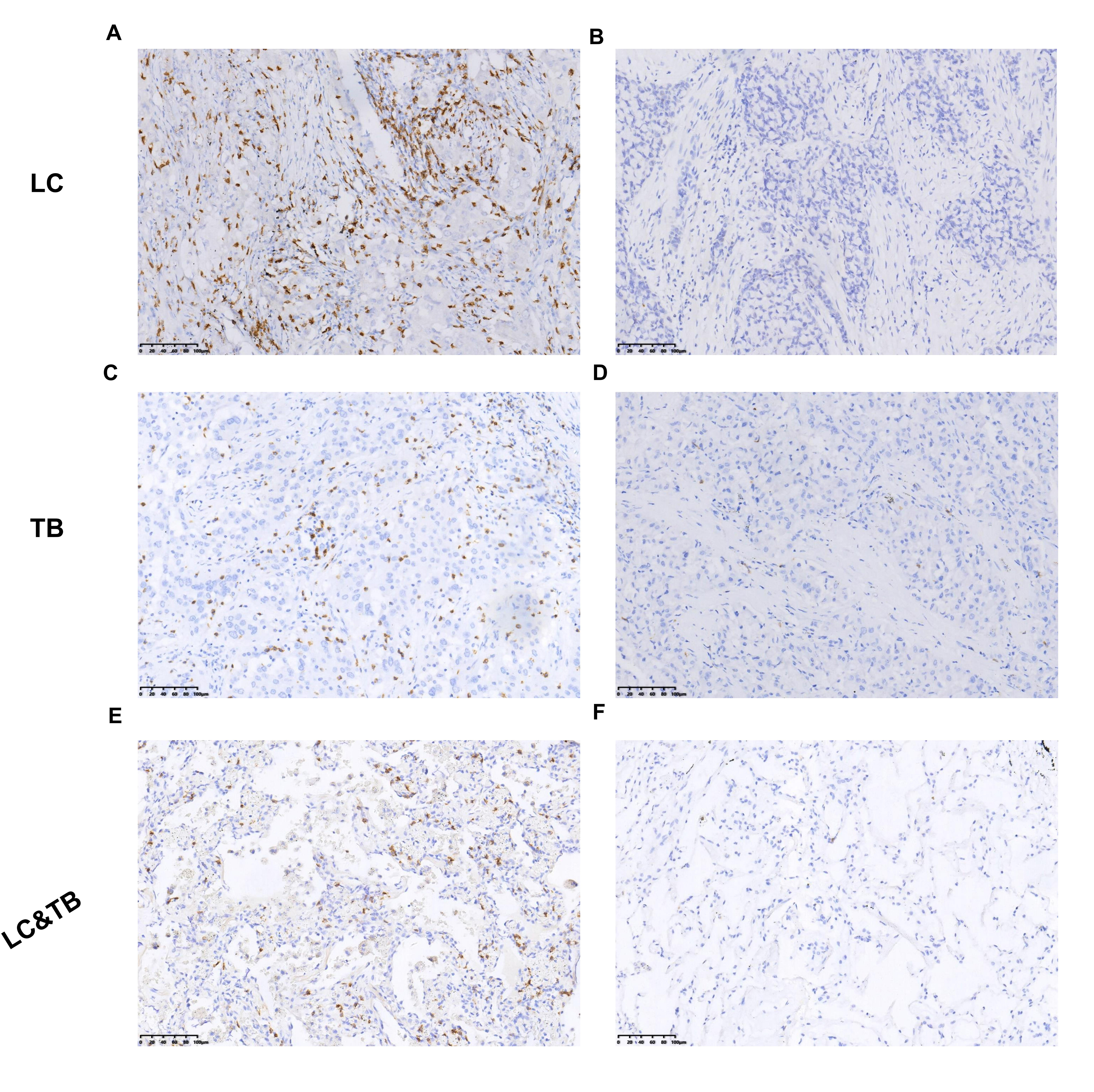
Preclinical studies have indicated that immune homeostasis in TB is regulated via immune checkpoint pathways, such as the PD-1/PD-L1 axis. For example, in animal models, the PD-1 pathway plays a key role in controlling excessive inflammation after MTB infection and regulating the resulting immune response (24). Furthermore, the PD-1/PD-L1 pathway suppresses the accumulation of CD4+ T-cells and IFN-γ production, which is an essential part of the immune response to TB. However, treatment that targets PD-1/PD-L1 may cause CD4 + T-cells to overproduce IFN-γ, which can aggravate TB or cause TB recurrence (24–35). Another study has indicated that a TB antigen can inhibit the Th1 immune response and promote LC metastasis via the PD-1/PD-L1 signalling pathway (36). However, there are limited data regarding the immune microenvironment in patients with LC and previous/current TB, which highlights the need for additional information regarding PD-L1 expression and TILs in the tumour and surrounding microenvironment. Our findings revealed that, relative to the LC group, the LC&TB group had significantly decreased PD-L1 expression and less CD3+/CD4+/CD8+ T-cell infiltration, which suggests that these patients have immunologically cold tumours in a noninflammatory microenvironment (Fig. 6A). To the best of our knowledge, this is the first comprehensive analysis of the tumour microenvironment landscape in LC&TB and LC.
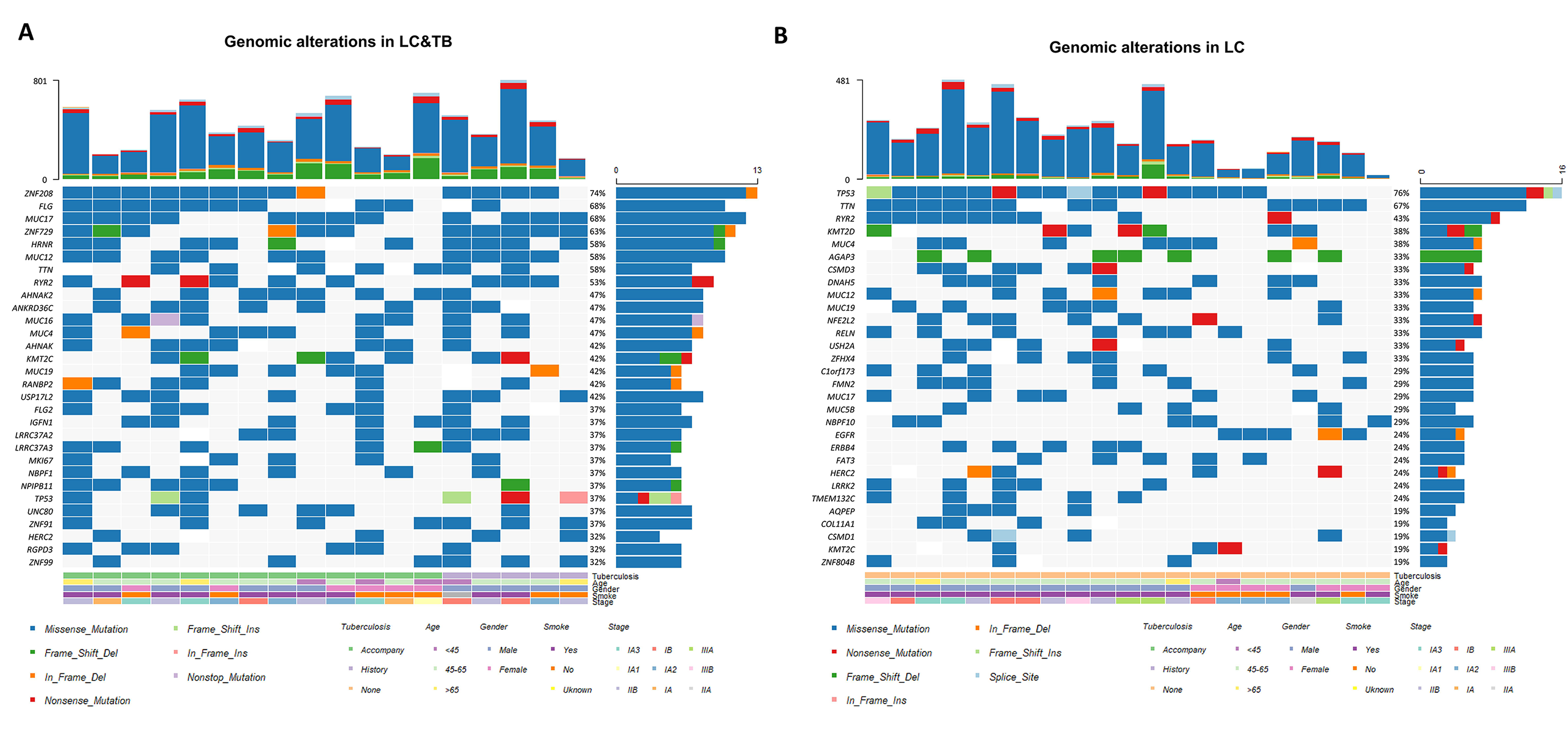
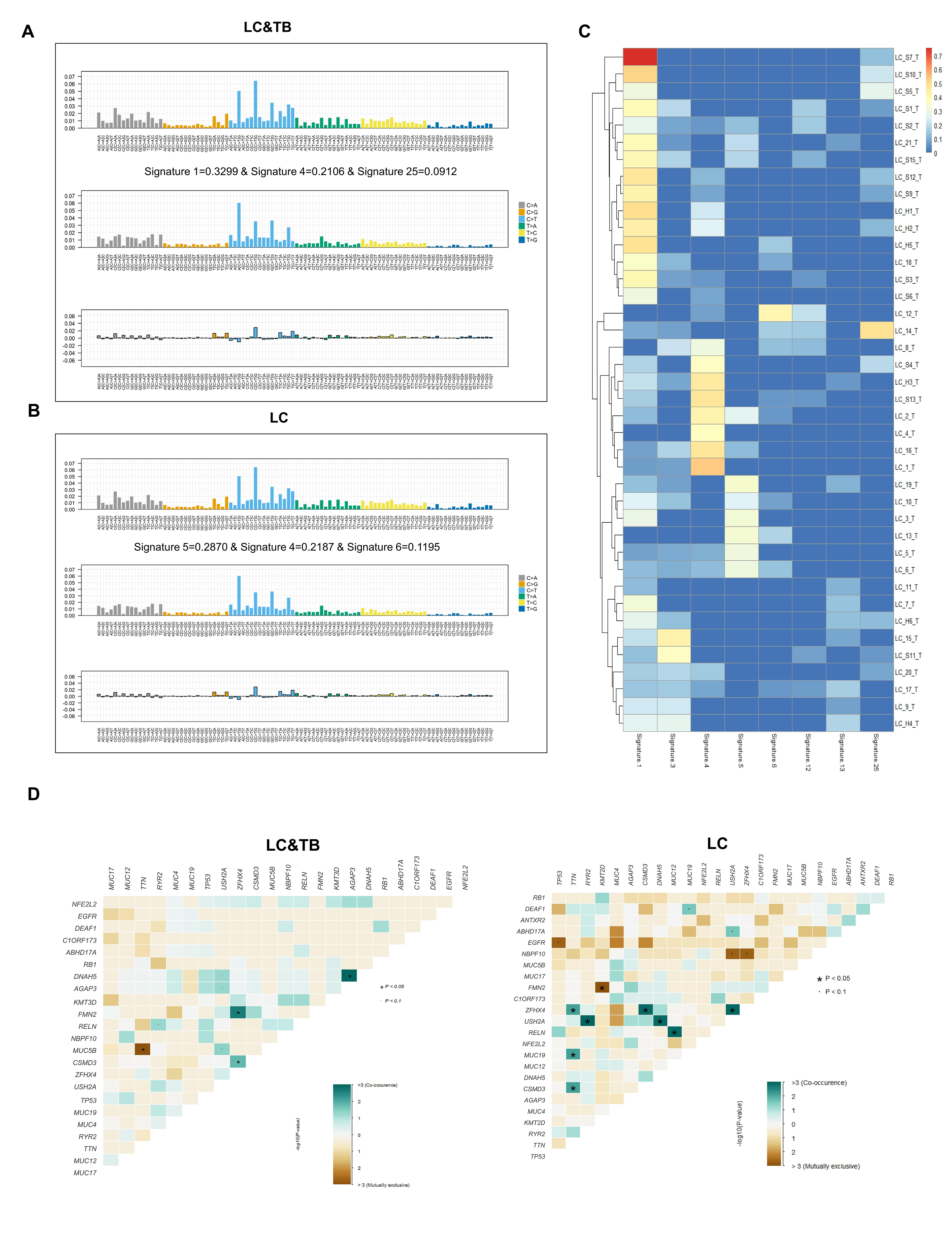
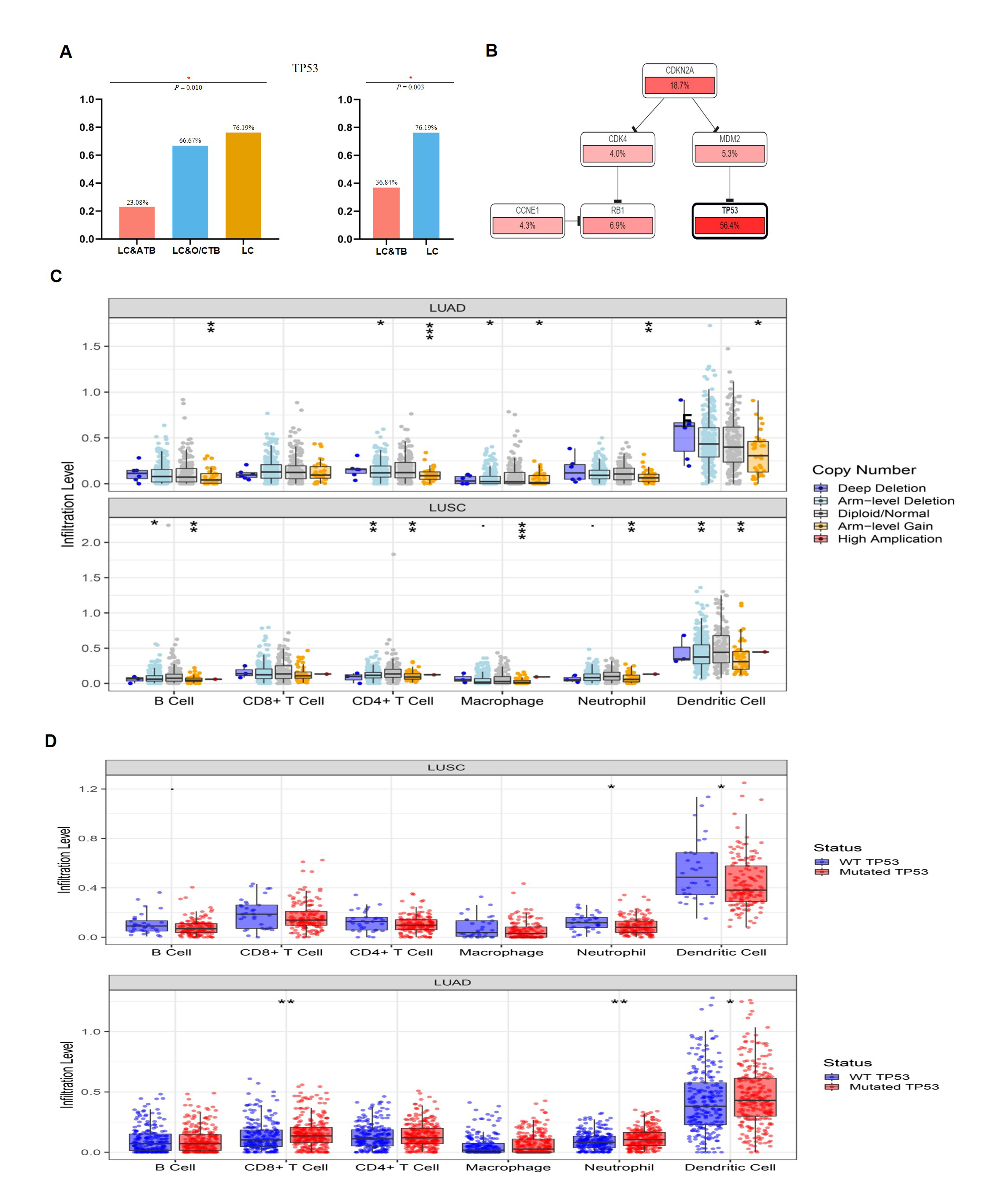
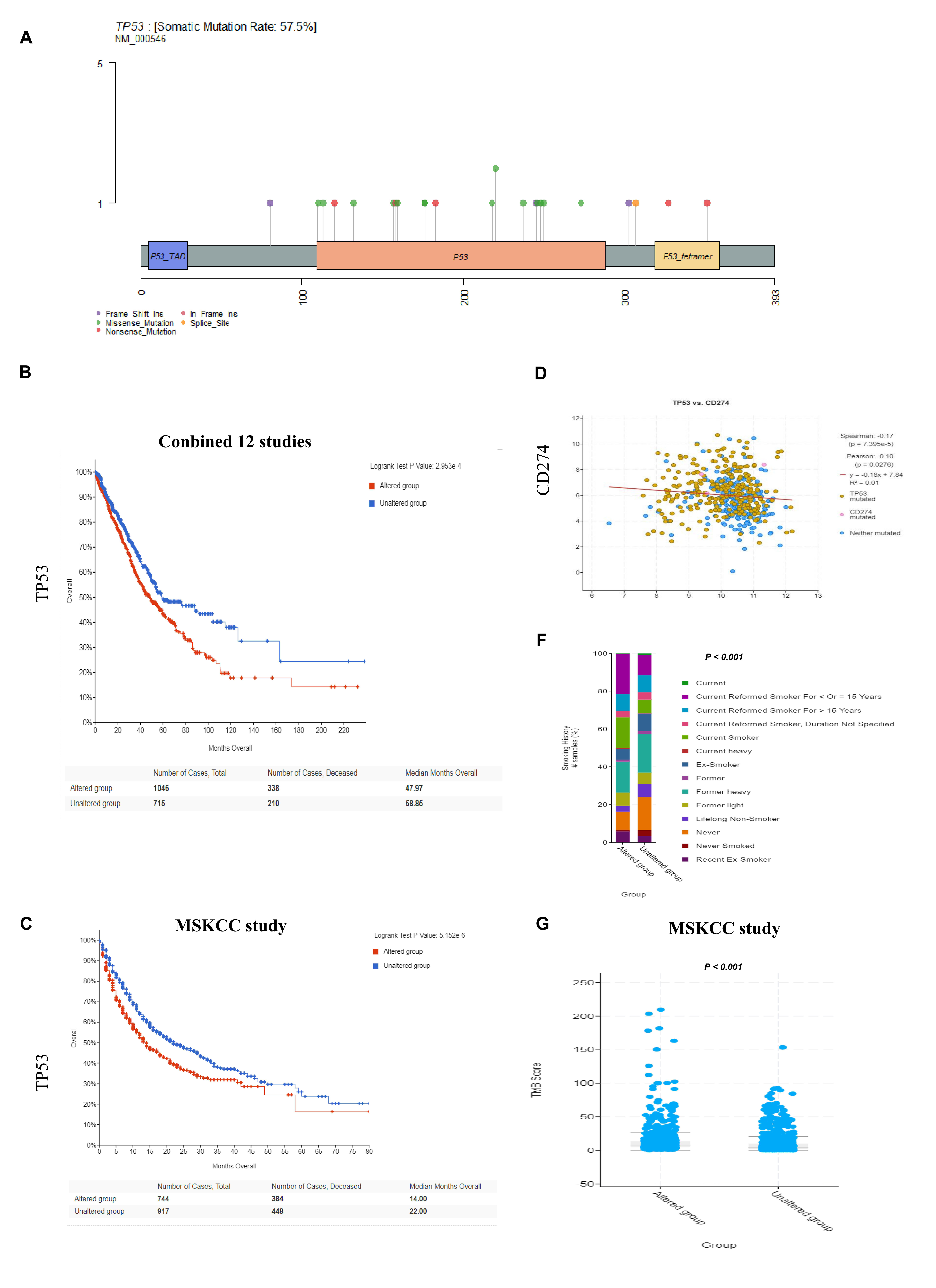
Previous studies have suggested that PD-L1 expression, TP53 mutation frequency, TMB (37), and HLA molecules may be biomarkers for predicting the response of LC to anti-PD-1/PD-L1 therapy (38–40), although these relationships remain controversial. In our study, the LC&TB group had a markedly higher TMB than the LC group, and the LC&TB group also had a significantly lower TP53 mutation frequency. The present study revealed that the LC&TB and LC groups only had heterozygous HLA I genotype, and thus we did not perform any additional analyses. Interestingly, we also observed that the number of nonsynonymous mutations was correlated with the number of neoantigens, which is consistent with previous research (41). Moreover, the TMB was significantly associated with smoking history among patients with LC&TB and LC alone, which also agrees with previously reported results (42). These results suggest that there are significant differences in terms of genomic alterations and mutation signatures between the LC&TB and LC groups.
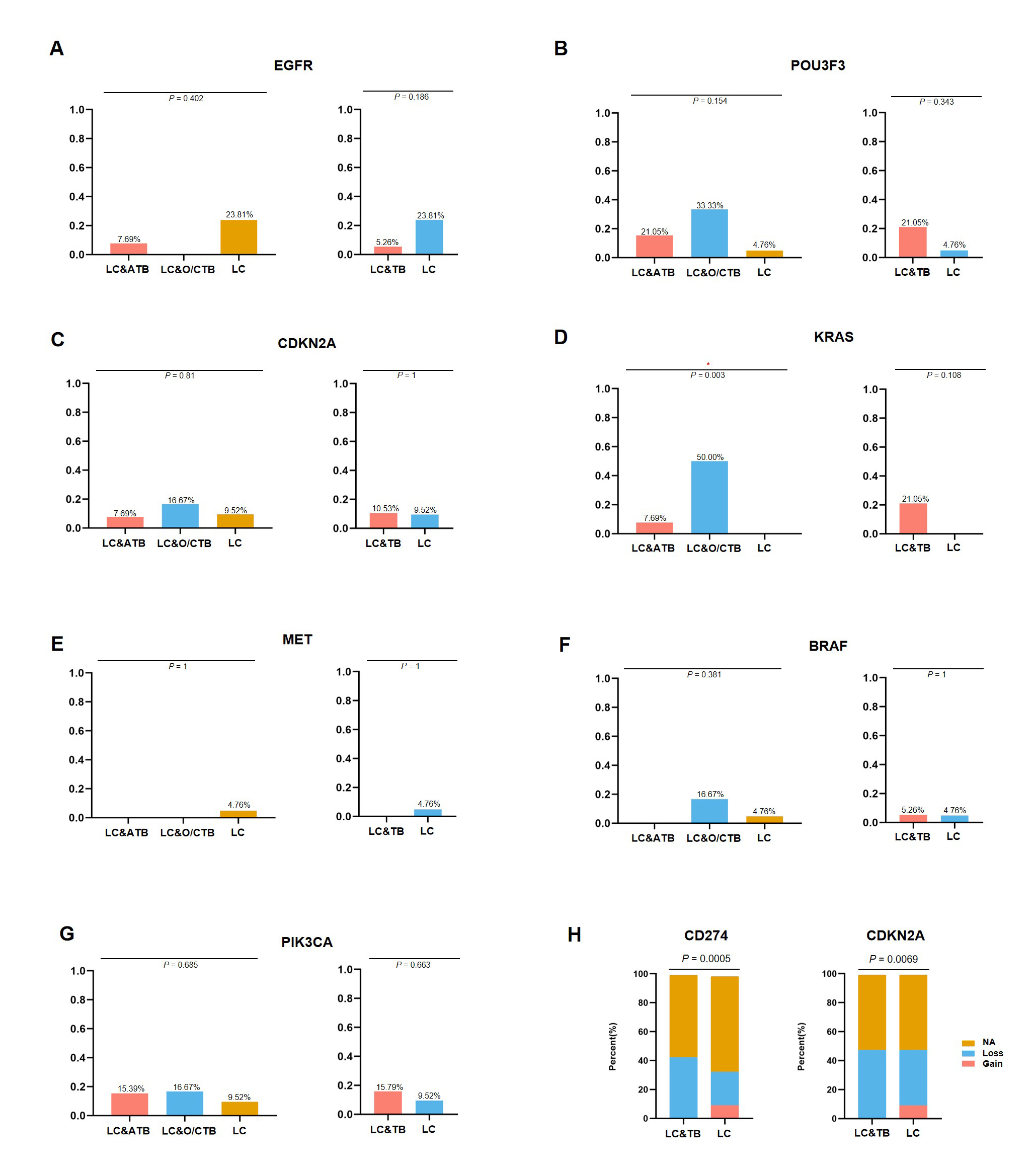
A few studies have attempted to identify potential driver genes in patients with LC who had a history of TB (43, 44). Adenocarcinoma was the main pathological type of LC among our patients with LC&TB, which is consistent with previously reported results (44). Although, patients with lung adenocarcinoma who have a history of lung scarring or TB are more likely to develop EGFR mutations, relative to patients with conventional lung cancer (44). Nevertheless, the present study failed to detect a significant difference between the LC&TB and LC groups in terms of the EGFR mutation frequencies.
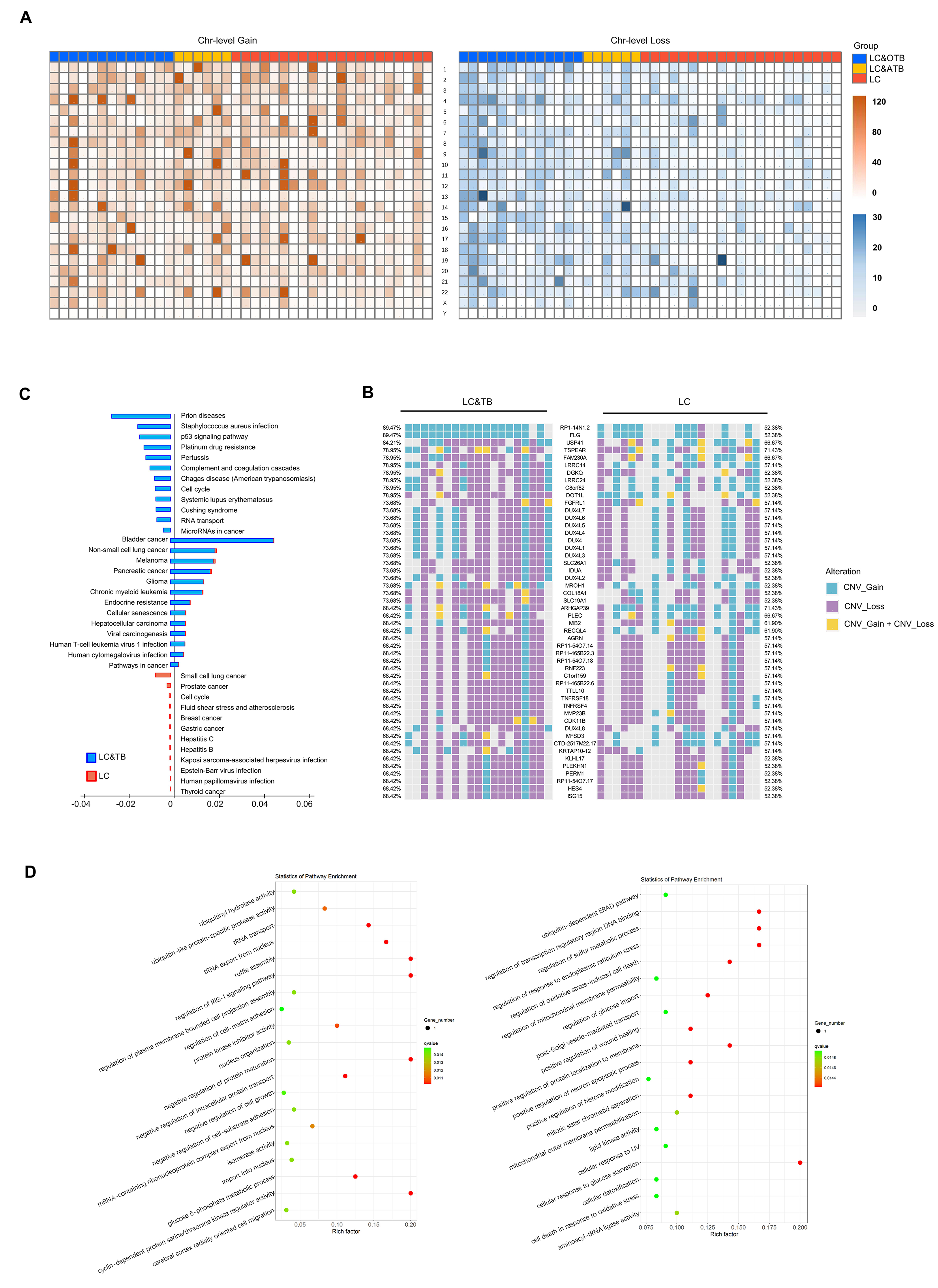
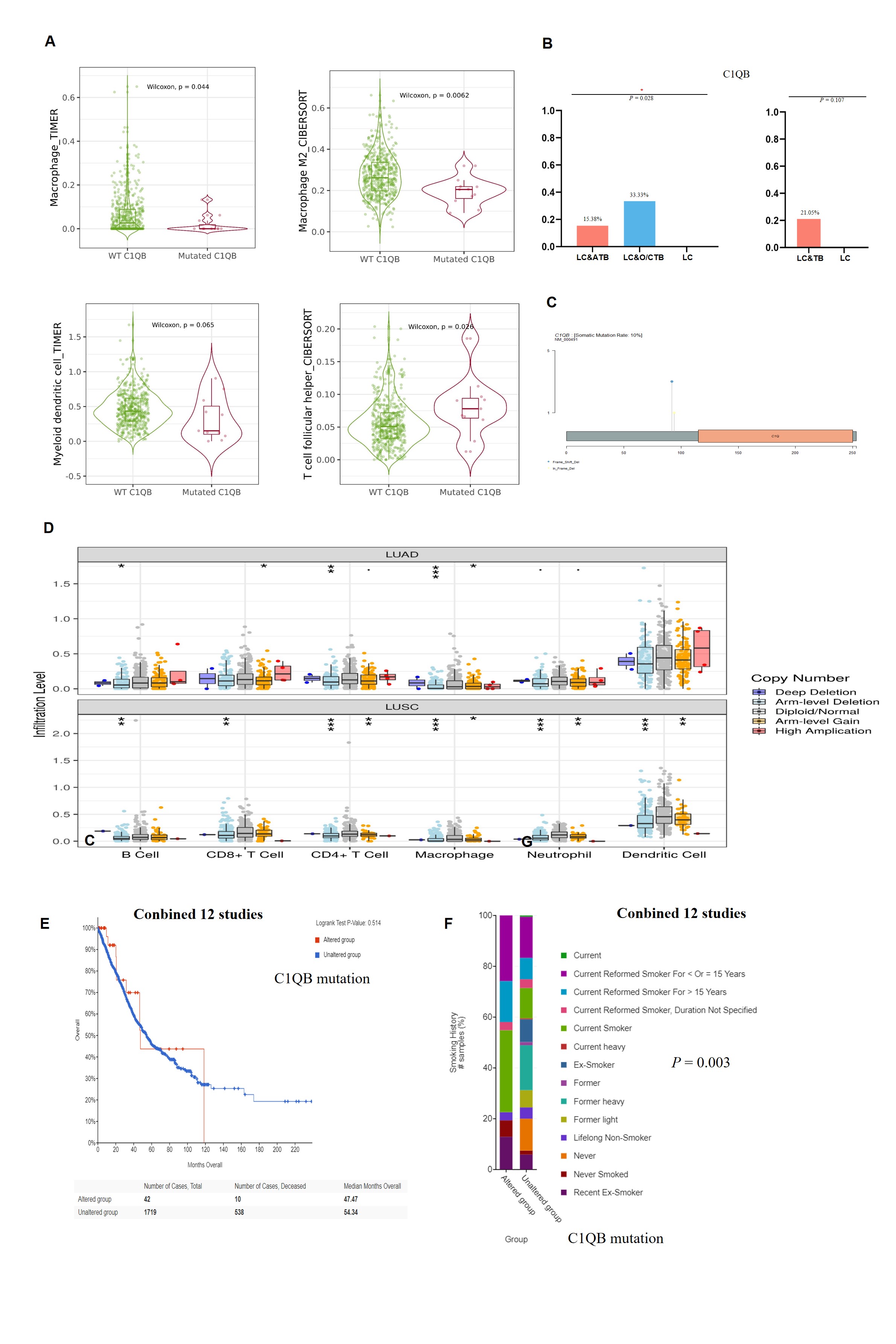
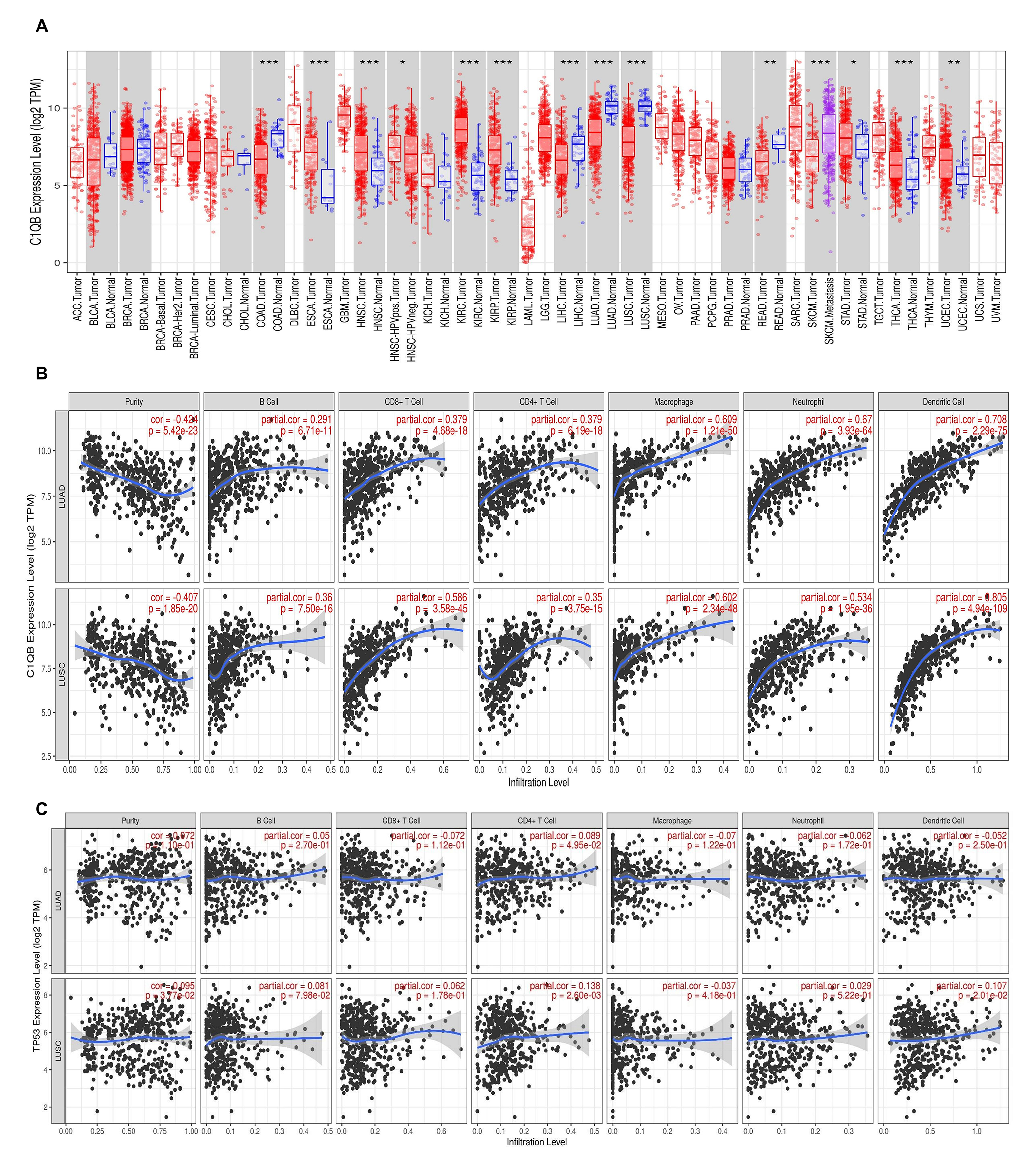
Notably, there were potential gene mutations differences between ATB and O/CTB in present study. For instance, The C1QB gene encodes the B-chain polypeptide of complement subcomponent C1q. The complement pathway is an important part of the immune system, and complement-mediated bacteriolysis and cytolysis are important mechanisms in the response to infection by pathogenic microorganisms. We observed that it was mutated at a higher rate among patients with LC&TB (5/19 cases) than among patients with LC alone (0/21 cases). In addition, bioinformatics analysis predicted that C1QB c.274_311del would have a substantial effect on protein expression. Furthermore, the C1QB gene may influence macrophage and dendritic cell infiltration of NSCLC. Therefore, C1QB may be a driver gene that leads to the developing of LC in patients with lung scarring or TB. In addition, KRAS mutation differences are highly significant between ATB and O/CTB (P = 0.003 in Supplementary Figure S6D), but when combined as a single group, there is no difference between LT&TB and LC alone. However, due to the limited sample size of these two groups, it is difficult to draw a clear conclusion. Further studies with large sample size and mechanism investigation are urgently needed.
Because a noninflammatory microenvironment we have observed in LC&TB and the possibility of reactivation/exacerbation of TB by anti-PD-1/PD-L1 therapy, which suggests that there is a need for new approaches to treating patients with LC&TB. We propose three potential strategies for treating patients with LC&TB (Fig. 6B). First, chimeric antigen receptor T-cell immunotherapy strategies that directly target CD8+ T-cells might help avoid overactivation of CD4+ T-cells. Second, dendritic cell vaccines could be developed for patients with LC&TB. Third, it might be useful to consider the possibility of drugs that target mutated C1QB.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}