Our study further supports the current evidence that Onchocerca cervipedis is not a single species, but a widely distributed species complex. We discovered that Onchocerca isolates found in Lake County, northern California, black flies belong to the same cryptic Onchocerca species previously found in Los Angeles County, southern California, black flies [17]. Corroborating the findings from southern California, Onchocerca DNA was detected in two black fly species: S. vittatum (s.l.) and S. tescorum [17] (Table 1). In addition, a third species of black fly was shown to carry the same cryptic Onchocerca DNA: S. clarum belonging to the “black” phenotype (Table 1).
Phylogenetic analyses of the nad5 gene demonstrates that the cryptic Onchocerca found in southern and northern California black flies (present study; [17]) and the equally cryptic Onchocerca isolate found in New York, northeastern USA, [16] represents one individual clade with little genetic divergence (Fig. 2). However, a definitive conclusion on whether the Californian isolates are conspecific with the New York isolates cannot yet be determined (Table 2; Fig. 3). Further studies targeting a multi-locus approach could help shed light on the exact phylogenetic relationships and taxonomic status of these geographically distant isolates. This notion is best exemplified by comparing the nad5 gene to the cox-1 gene, which appears to exhibit greater diversity within the cryptic Onchocerca isolates [17]. In addition, at this stage, it is not possible to conclude that the cryptic species present in northern California belongs to the originally described O. cervipedis. In the original description of the species by Wehr and Dikmans [28], the authors used specimens from two different locations and from at least two different hosts, including O. virginianus and O. hemionus from Montana, USA and O. hemionus from British Columbia, Canada. To further elucidate this taxonomic conundrum, isolates from these hosts and locations should be collected, morphologically re-evaluated, molecularly characterized, and subsequently compared to these many isolates within the O. cervipedis complex.
Molecular screening and putative vectors of cryptic Onchocerca isolates
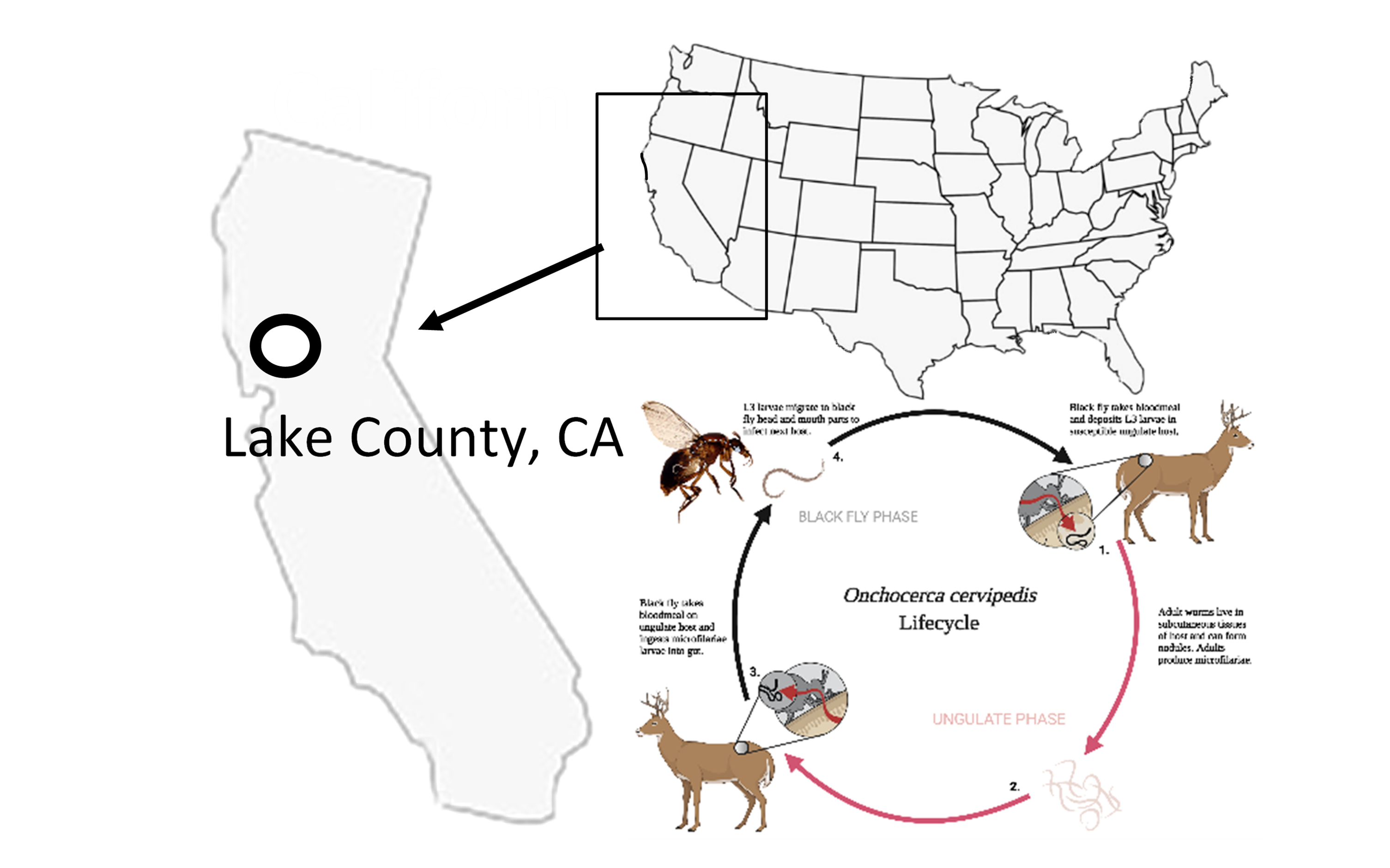
The finding of cryptic Onchocerca DNA through molecular screening of arthropod vectors (i.e., xenomonitoring) provides a straightforward approach to understanding more about parasite biodiversity, geographic distribution, and putative vector associations. In addition, vector screening allows researchers to not solely rely on labor-intensive measures such as post-mortem examination of host carcasses for adult worms [28] or an unstandardized technique to obtain microfilariae [29, 30], given the potential variable predilection sites of adults and microfilariae of the species within the O. cervipedis complex. For instance, while evidence indicates that microfilariae could be present near the skin of the ear [29, 31–33], the true predilection site of O. cervipedis species complex microfilariae does not have clear a consensus [34, 35] and what, if any, variation exists among ungulate host species. Moreover, the utilization of xenomonitoring of North American parasites allows for concurrent monitoring of other similar Onchocerca species, such as the zoonotic O. lupi, that are of current public health concern [36].
Comparable to Verocai et al. [17], our results showed the positive rate for Onchocerca DNA was low in the black fly populations. Indeed, this low prevalence is an expected result of pool sampling, as prior evidence has shown that pool samples are highly sensitive to low prevalence of pathogens in vector species [37]. Furthermore, other closely related filarial nematode studies revealed similarly low positive prevalence rates of O. lupi in southern California [36], O. volvulus in Africa [38, 39], and Wuchereria bancrofti (Cobbold, 1877) in American Samoa and Guinea [40–42].
Our study also provided evidence for an additional species of black fly as a probable vector of this Onchocerca species. According to the literature, S. clarum has been reported to feed on a variety of mammals (horses, cattle, rabbits, and humans) and birds [43, 44]. The finding of DNA of an Onchocerca species likely associated with a cervid host(s) suggest that these mammals may serve as a blood source for this dipteran, similar to that of S. tescorum and S. vittatum, as suggested by Verocai et al. [17, 20]. However, S. clarum is restricted to the Central Valley California region near the present study site of Lake County [20]. Similarly, S. tescorum has been reported with a limited range, spanning only California and Arizona [20, 23]. This means that even if these two vectors are competent host for this Onchocerca species, they would only contribute to the transmission within their more restricted distribution. In contrast, species within the S. vittatum complex which includes S. tribulatum, have a widespread distribution across North America, including both California and New York [23]. We postulate that it could play an epidemiologically relevant vector role for this Onchocerca isolate found in northern and southern California (present study; [17, 36]) and possibly New York [16].
Definitive hosts of cryptic Onchocerca isolates
The recent discoveries of at least two or more genetic Onchocerca isolates in North America likely associated with at least three of the cervid hosts (i.e., mule deer, white-tailed deer, and moose) raise many questions regarding Onchocerca-host assemblages. Of these three cervid hosts, only the mule deer’s range encompasses southern California, including Los Angeles county [45–47]. Thus, it was reasonably hypothesized that the mule deer could be the putative host to the Onchocerca isolate from southern California [17]. Lake County also includes the range of the mule deer [45], however, unlike southern California, Lake County is also home to the Californian Tule elk or Cervus elaphus nannodes Merriam, 1905 [48]. This elk subspecies was hunted to near extinction in the late 1800s, and now has a thriving population in California. According to most recent data, about 6,000 Tule elk populate California, including many herds that live near the Lake County region of northern California where black flies were sampled for this current study [48–50]. While there was no blood meal analysis completed, it is possible that these cervids could be a blood meal source for black flies and consequently be a potential host to the O. cervipedis species complex [7]. Ideally, adult worms or microfilariae should be sampled from necropsied elk hosts and molecularly analyzed to confirm its definitive host status.
Species within O. cervipedis complex have been reported in a variety of locations across North America in the six ungulate hosts: pronghorn from Idaho [8]; moose from Alaska, Alberta, British Columbia, and Northwest Territories [11, 13–15, 35]; elk from Montana [7]; mule deer from Arizona, California, Montana, Utah, Wyoming, and British Columbia [3, 4, 6, 7, 9, 17, 28, 29, 33, 34, 51–56]; white-tailed deer from Arizona, Missouri, Montana, New York, Oregon, Pennsylvania, British Columbia, and also from Costa Rica [4, 5, 7, 12, 16, 28, 29, 31, 32, 52, 57–59]; and caribou from Alaska and British Columbia [10, 14]. Additional records from Odocoileus from Colorado, Idaho, and Montana, were reported as “deer”, without species designation [60–64]. Therefore, it can be inferred that sample collection should begin in these reported locations and include all six ungulate hosts when obtaining biological samples. Recovery of nematodes from necropsy, with subsequent morphological and DNA identification, will confirm parasitic infection of a definitive host and aid in interpreting the distribution of cryptic Onchocerca isolates.
Evolutionary history and ecological considerations of cryptic Onchocerca isolates
Currently, it is hypothesized that the two, and possibly more, known Onchocerca species (i.e., O. cervipedis sensu Verocai et al. [14] and the clade comprising the Californian and New York isolates [16, 17]) are the result of independent expansion events from Palearctic ungulates hosts colonizing from across the Bering Landbridge into the Nearctic [65–67]. It is currently unknown if the finding of at least two Onchocerca species within the complex is the result of a small, incomplete sampling of a larger species complex or the true representation of diversity in North America. Nevertheless, there is substantial evidence from eastern Asia for prior underestimation of Onchocerca species diversity and richness. For instance, Onchocerca suzukii Yagi, Bain and Shoho, 1994, Onchocerca eberhardi Uni et al., 2007, and Onchocerca takaokai Uni, Fukuda and Bain, 2015 have been recently described from wild ungulates of Japan [68–70]. Furthermore, Onchocerca borneensis Uni, Mat Udin & Takaoka, 2020 [71], was described in bearded pigs of Borneo with additional molecular evidence suggesting two closely related parasites, Onchocerca dewittei Bain, Ramachandran, Petter & Mak, 1977 and Onchocerca japonica Uni, Bain and Takaoka, 2001, which were considered subspecies of the former were, in fact, separate species [71]. Indeed, it is feasible that the North American Onchocerca species complex, which much is still unknown about, could comprise undescribed Onchocerca diversity, similar to the pattern that we have witnessed in Asian suids and ungulates. Moreover, host-parasite biogeography appears to play a critical role in Onchocerca diversification. As noted by Uni et al. [71], O. borneensis and O. dewittei infect Sus barbatus Müller and Sus scrofa vittatus Boie in the Indomalayan region, but O. japonica and O. dewittei infect different subspecies of the same host species in the Palearctic and Indomalayan regions. Thus, when re-evaluating Onchocerca in the North American landscape, collecting specimens from sympatric and allopatric host ranges may yield more complete information about parasitic diversity.
{kind=link}