Clinical implications of abnormally expressed circASCC3 in NSCLC
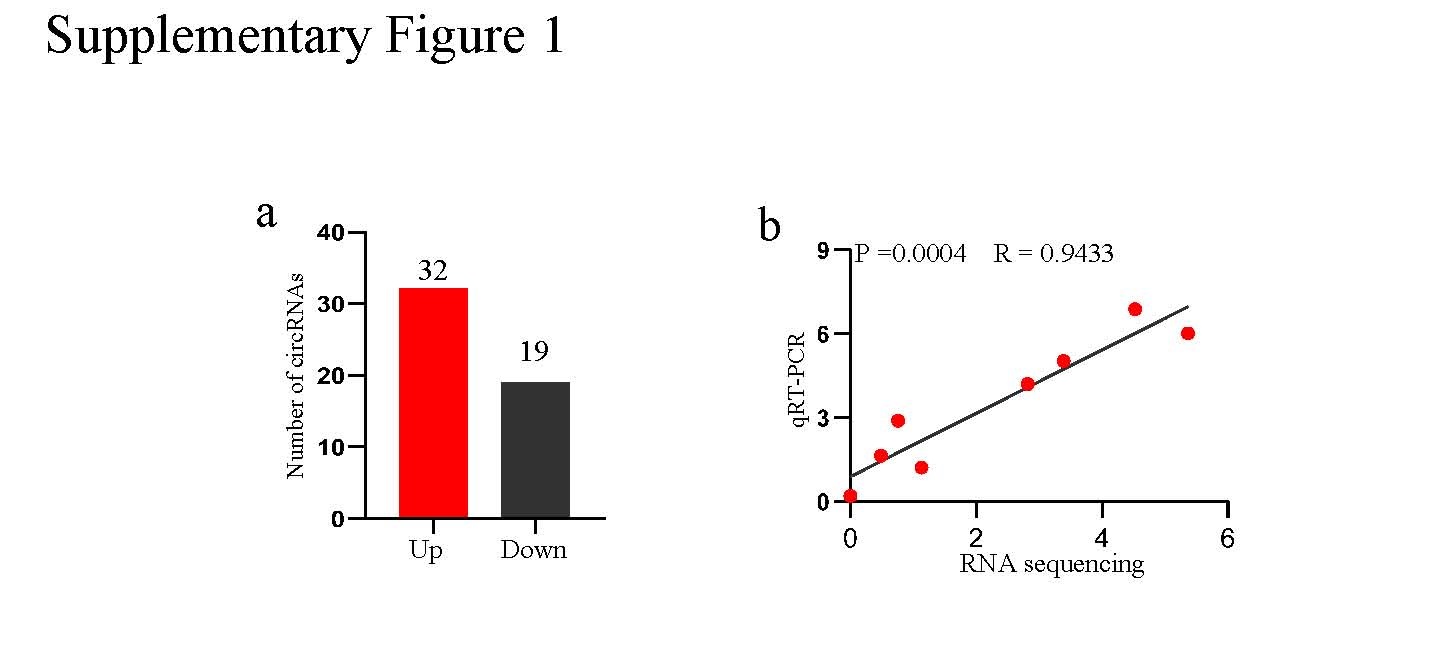
First, we conducted circRNA sequencing to explore differentially expressed circRNAs between NSCLC and matched normal tissues. A total of 32 circRNAs were obviously upregulated, while 19 circRNAs were downregulated in NSCLC tissues (Supplementary Fig. 1a). Among these differentially expressed circRNAs, the expression of circASCC3 (hsa_circ_0077495) was the most significantly different, and it was consistently upregulated in 4 NSCLC tissues; moreover, the back splice site of circASCC3 was confirmed by Sanger sequencing with divergent primers (Fig. 1a, b). In addition, the detection of circASCC3 in the 4 pairs of NSCLC tissues and matched nontumor tissues by qRT-PCR further confirmed the reliability of circRNA sequencing (Supplementary Fig. 1b). The PCR results showed that circASCC3 could be detected by divergent primers in cDNA reverse transcribed by random hexamers rather than oligo dT primers (Fig. 1c). Further experiments with RNase R exonuclease confirmed the covalent closed-loop structure of circASCC3 (Fig. 1d). Since circASCC3 is located on chromosome 6q13, a location known to carry chromosomal aberrations that contribute to tumor progression, we speculated that circASCC3 might potentially regulate the biological function of NSCLC. Next, circASCC3 expression was detected in 35 NSCLC tissues and matched normal tissues using qRT-PCR, and the results showed that circASCC3 was highly expressed in 23/35 NSCLC tissues (Fig. 1e).
To identify the clinical relevance of circASCC3, the expression of circASCC3 was evaluated in 188 pairs of NSCLC tissues and matched peritumor tissues in a TMA by ISH. The expression of circASCC3 was obviously elevated in NSCLC tissues (Fig. 1f). The results of the analysis of circASCC3 expression and clinical characteristics of 188 NSCLC patients are presented in Table 1. The results suggested that a high level of circASCC3 was associated with a larger tumor size, the appearance of lymph node metastasis and an advanced TNM stage. Therefore, we further explored the potential impact of circASCC3 expression on the prognosis of NSCLC patients. Consistently, the expression of circASCC3 was correlated with recurrence and OS. Patients with high expression of circASCC3 had an obviously higher postoperative recurrence rate and shorter OS (Fig. 1g, h). Moreover, the Cox regression analysis showed that the expression of circASCC3 was an independent predictor for recurrence and OS (Tables 2, 3). In short, these results reveal that circASCC3 may exert an influence on the progression of NSCLC.
circASCC3 enhances invasion, metastasis and proliferation in NSCLC
To further investigate the biological function of circASCC3 in NSCLC, the expression of circASCC3 was measured in 4 NSCLC cell lines, A549, H1299, H460, and H1703, via qRT-PCR. CircASCC3 expression was highest in A549 cells and lowest in H1299 cells (Fig. 2a). Therefore, we designed and synthetized two lentiviral vectors for the knockdown and overexpression of circASCC3 to establish two stable cell lines (A549-shcircASCC3 and H1299-circASCC3, respectively) (Fig. 2b). Matrigel Transwell assays showed that the invasive potential was significantly enhanced after overexpression of circASCC3 (Fig. 2c). Consistently, the wound healing assays also suggested a similar tendency; the wound healing ability was reduced in A549-shcircASCC3 cells but enhanced in H1299-circASCC3 cells (Fig. 2d). In terms of the proliferation ability, the colony formation assay and CCK-8 assay showed that overexpression of circASCC3 enhanced the proliferation of NSCLC cells, while knockdown of circASCC3 led to the opposite effect (Fig. 2e, f). Furthermore, we verified the in vitro results in nude mice. The results showed that the mice implanted with H1299-circASCC3 cells had a larger tumor burden and were more prone to lung metastasis than those implanted with control cells (Fig. 2g, h). Thus, these results indicate that circASCC3 promotes a malignant phenotype in NSCLC.
circASCC3 functions as a sponge for miR-432-5p
Serving as a miRNA sponge has been identified as an important function of circRNAs. Thus, we hypothesized that circASCC3 might promote the progression of NSCLC by binding to miRNAs. We found 34 miRNAs that were potential targets for circASCC3 in StarBase. To further screen the miRNAs that were possibly sponged by circASCC3, a circASCC3 probe was designed to purify the circASCC3-related miRNAs using circRIP in A549 cells, and then, the predicted miRNAs were evaluated by RT-qPCR. The results showed an obvious enrichment of miR-432-5p in complex with circASCC3 in contrast to the negative control, which suggested that miR-432-5p might be one of the critical miRNAs interacting with circASCC3 to regulate its biological function in NSCLC (Fig. 3a). To further prove that circASCC3 serves as a sponge for miR-432-5p, a dual-luciferase reporter gene assay was performed in HEK-293T cells. The wild-type (WT) and mutant circASCC3 sequences were copied into the vector pLG3 (Fig. 3b). The miR-432-5p mimic obviously weakened the luciferase activity of WT circASCC3 but not mutant circASCC3 (Fig. 3c). In addition, the pulldown assay with a biotinylated miR-432-5p probe resulted in significant enrichment of circASCC3 in A549 and H460 cells (Fig. 3d). Moreover, further knockdown of miR-432-5p with miR-432-5p inhibitor rescued the migration, invasion and proliferation ability of A549-shcircASCC3 and H460-shcircASCC3 cells (Fig. 3e-h). In FISH analysis, circASCC3 and miR-432-5p were colocalized in the cytoplasm of NSCLC cells (Fig. 3i). Taken together, these findings indicate that circASCC3 sponges miR-432-5p in NSCLC.
Sponging of miR-432-5p by circASCC3 enhances the expression of C5a
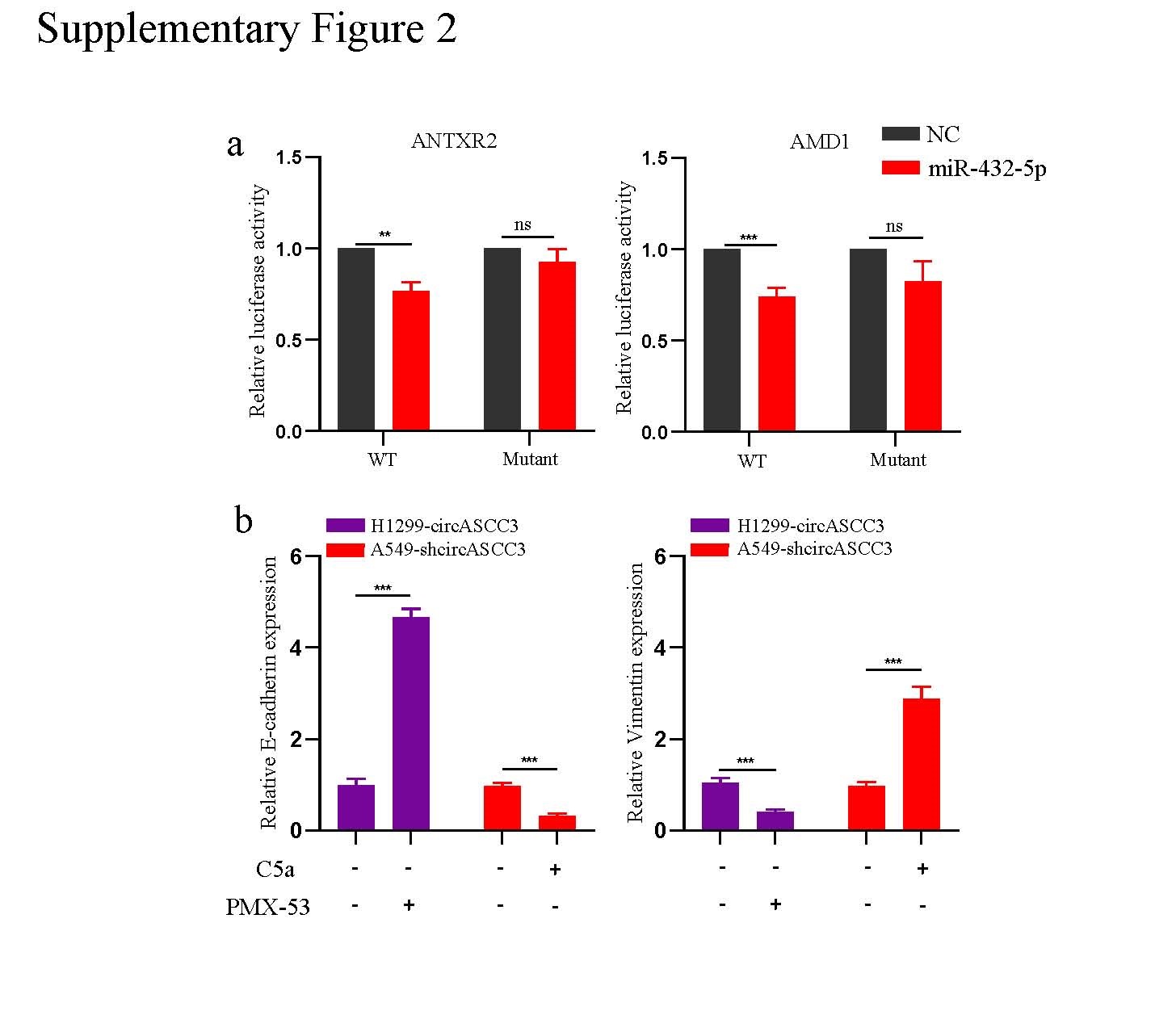
To further identify the downstream mechanism of circASCC3 in promoting the progression of NSCLC, we performed RNA-seq to explore the differentially expressed genes between H1299-control and H1299-circASCC3 cells and discovered the upregulated expression of 84 genes and downregulated expression of 65 genes in H1299-circASCC3 cells (Fig. 4a). Reactome analysis suggested that these genes showed significant enrichment of processes involved in the immune system. KEGG analysis showed enrichment of pathways in cancer, cell cycle, PI3K-Akt signaling pathway, T cell receptor signaling pathway and leukocyte transendothelial migration, and PANTHER analysis showed enrichment of the TGF-beta signaling pathway, EGF receptor signaling pathway and Wnt signaling pathway (Fig. 4b). We also predicted the potential targets of miR-432-5p by overlapping the 149 differentially expressed genes identified from the RNA-seq experiments with the predicted targets from TargetScan and the miR-432-5p pulldown experiment results, and the results showed that ZNF579, STK16, C5, ANTXR2, ANKRD42 and AMD1 were potential candidate genes (Fig. 4c). Since previous studies have reported an oncogenic role of C5, ANTXR2 and AMD1[16–18], we assessed these three genes in separate dual luciferase assays to determine whether they are targets of miR-432-5p. The luciferase activity was dramatically reduced in HEK-293T cells carrying the C5 WT sequence compared to HEK-293T cells carrying the mutant sequence of C5 (Fig. 4d, e, Supplementary Fig. 2a). Moreover, the expression of C5 mRNA was decreased in A549 cells with knockdown of circASCC3 and increased in H1299 cells after overexpression of circASCC3 (Fig. 4f). Previous studies have identified C5a (a hydrolysate of C5) as an important mediator in the progression of cancer[19, 20]. Thus, we also detected the expression of C5a in the supernatant using ELISAs, and the results showed that the concentrations of C5a were increased in the supernatant of H1299 cells after forced expression of circASCC3 and decreased in the supernatant of A549 cells with knockdown of circASCC3 compared with the relative values in the supernatant of control cells (Fig. 4g). To further identify the role of circASCC3/C5a in NSCLC cells, we stimulated A549-shcircASCC3 cells with C5a (50 ng/ml) and inhibited the C5a/C5aR axis in H1299-circASCC3 cells with the C5aR inhibitor PMX-53 (20 nM). The Matrigel Transwell assay showed enhanced invasion after C5a stimulation, while PMX-53 treatment had the opposite effect (Fig. 4h). In addition, the relationship between circASCC3 and C5 was evaluated in 30 NSCLC patient tissues using qRT-PCR. The results showed that the expression of C5 was positively correlated with that of circASCC3 (Fig. 4i). Previously, we discovered that the C5a/C5aR axis was involved in the progression of NSCLC through epithelial–mesenchymal transition (EMT)[21]. Here, we identified the EMT phenotype in H1299-circASCC3- and C5a-induced A549-shcircASCC3 cells, and this phenotype could be reversed by PMX-53 (Fig. 4j, Supplementary Fig. 2b). Collectively, these results confirm the contribution of the circASCC3/miR-432-5p/C5a axis to the malignant phenotype of NSCLC cells.
circASCC3 induces an immunosuppressive phenotype of macrophages through C5a and further suppresses CD8+ T cell function via PD-L1
Since C5a has been identified as a critical regulator in tumor-associated macrophages (TAMs) and is downstream of circASCC3 in NSCLC[22], we speculated that circASCC3 may have a potential impact on the TME. Therefore, we established an LLC-circASCC3 subcutaneous xenograft model in C57BL/6 mice and further analyzed the immune profile of the tumor tissues by flow cytometry. The results showed that the infiltration of CD8+ T cells was dramatically reduced, while no obvious change in other subpopulations was observed (Fig. 5a). To further verify the in vivo results, we also detected the relationship between circASCC3 and CD8+ T cells in a TMA containing 188 NSCLC tissues by IHC (Fig. 5b). The results showed that the expression of circASCC3 was negatively correlated with CD8+ T cells, which was in line with the in vivo results. To identify the impact of circASCC3 on CD8+ T cells, we isolated CD8+ T cells from 5 healthy donors and cultured them with supernatants from H1299-circASCC3/control cells. However, the overexpression of circASCC3 in NSCLC cells did not impair the cytotoxic function or induce the apoptosis of CD8+ T cells, but the C5a level in the supernatant of H1299-circASCC3 cells was significantly upregulated (Fig. 5c-e). Thus, the results suggest that there is a mediator between circASCC3 and CD8+ T cells.
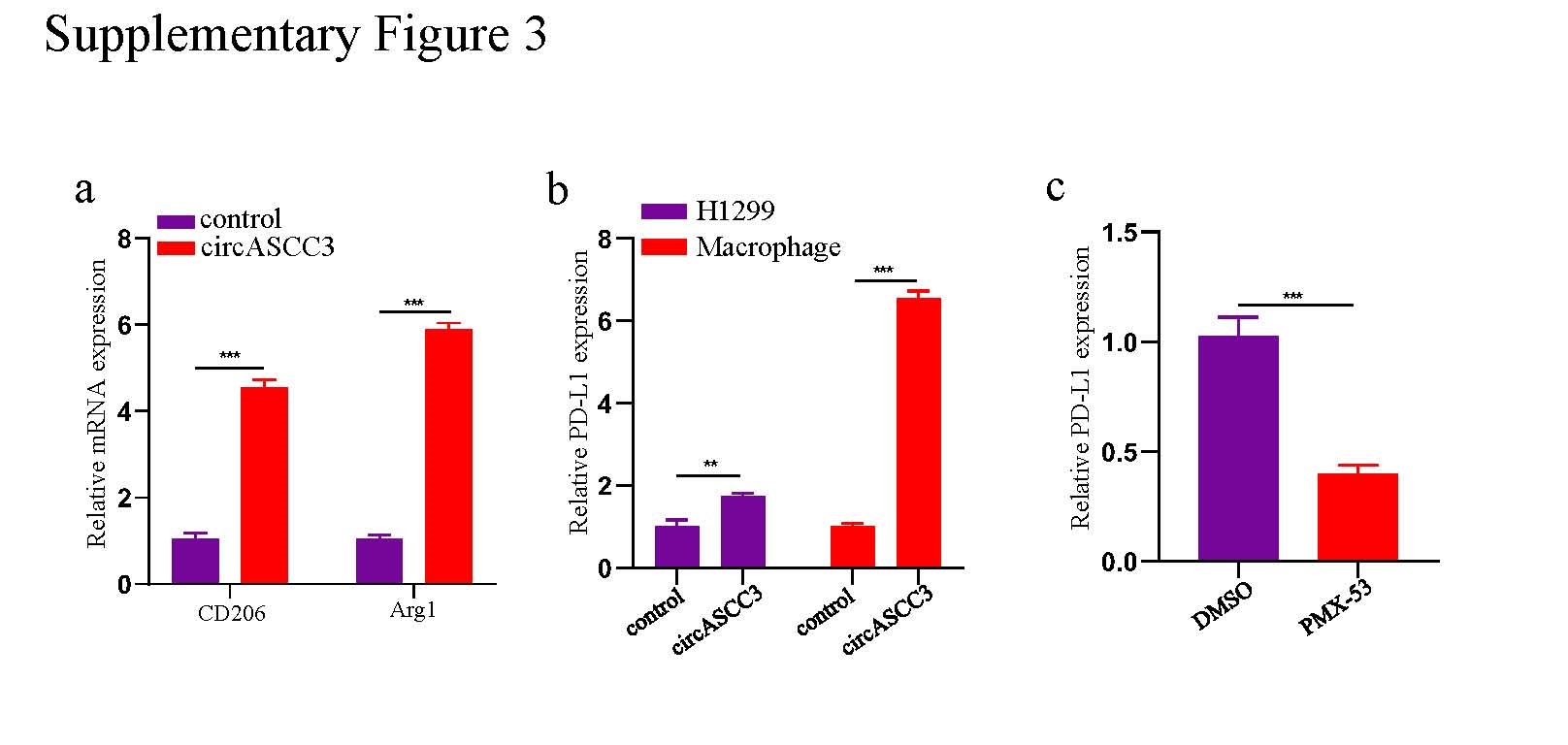
Since macrophages are a critical target of C5a and M2-type macrophages exert immune inhibition[23, 24], we established a 48-h coculture system of H1299-circASCC3 cells and PMA-stimulated THP-1 cells and further cultured CD8+ T cells with the supernatant from the coculture system for 72 h. Flow cytometry analysis showed that the macrophages cocultured with H1299-circASCC3 cells exhibited a CD163-high phenotype, which was not seen for macrophages cocultured with H1299-control cells (Fig. 5f). In addition, the qRT-PCR results showed significantly increased mRNA levels of CD163 and other M2 markers, such as CD206 and Arg-1, in macrophages cocultured with H1299-circASCC3 cells (Fig. 5g, Supplementary Fig. 3a). Moreover, the cytotoxic function of CD8+ T cells was obviously impaired, with a remarkable decline in IFN-γ and GZMB in the supernatant (Fig. 5h). The apoptosis of CD8+ T cells was also significantly elevated (Fig. 5i). The IHC staining of CD163 in the TMA containing 188 NSCLC tissues revealed that the infiltration of CD163+ macrophages was positively correlated with the expression of circASCC3 (Fig. 5j).
To determine whether there were differences in the levels of secreted cytokines in the supernatant of the circASCC3 coculture system, the levels of secreted cytokines in supernatants from the H1299-control cell/macrophage and H1299-circASCC3 cell/macrophage coculture systems were detected by a Luminex Human Cytokine Antibody Panel. The levels of IL-2 and IL-12 were significantly reduced, while those of M-CSF, IL-6, IL-8, PD-L1, TNF-a and VEGF were obviously increased in the H1299-circASCC3 cell/macrophage coculture system (Fig. 5k). Considering the critical role of the PD-L1/PD-1 axis in CD8+ T cell dysfunction and immune evasion, we detected the expression of PD-L1 in macrophages and H1299-circASCC3 cells in the coculture system by western blot and qRT-PCR. The results showed that circASCC3 overexpression drastically elevated the expression of PD-L1 in macrophages, while the expression of PD-L1 was only mildly increased in H1299 cells (Fig. 5l, Supplementary Fig. 3b). In addition, the impaired function of CD8+ T cells could be rescued when the C5aR inhibitor PMX-53 was introduced into the circASCC3 coculture system or when CD8+ T cells were cultured with the PD-L1 inhibitor durvalumab and the supernatant from the coculture system (Fig. 5m). The apoptosis of CD8+ T cells was also inhibited by these two agents (Fig. 5n). Notably, PMX-53 reversed M2-like polarization and reduced PD-L1 expression in macrophages cocultured with H1299-circASCC3 cells (Fig. 5o, p, Supplementary Fig. 3c). In summary, these results indicate that circASCC3 promotes an immunosuppressive phenotype of macrophages through the upregulation of C5a, which further induces the upregulation of PD-L1 and the dysfunction of CD8+ T cells.
PMX-53 improves the effects of anti-PD-1 immunotherapy in a xenograft model with circASCC3 overexpression
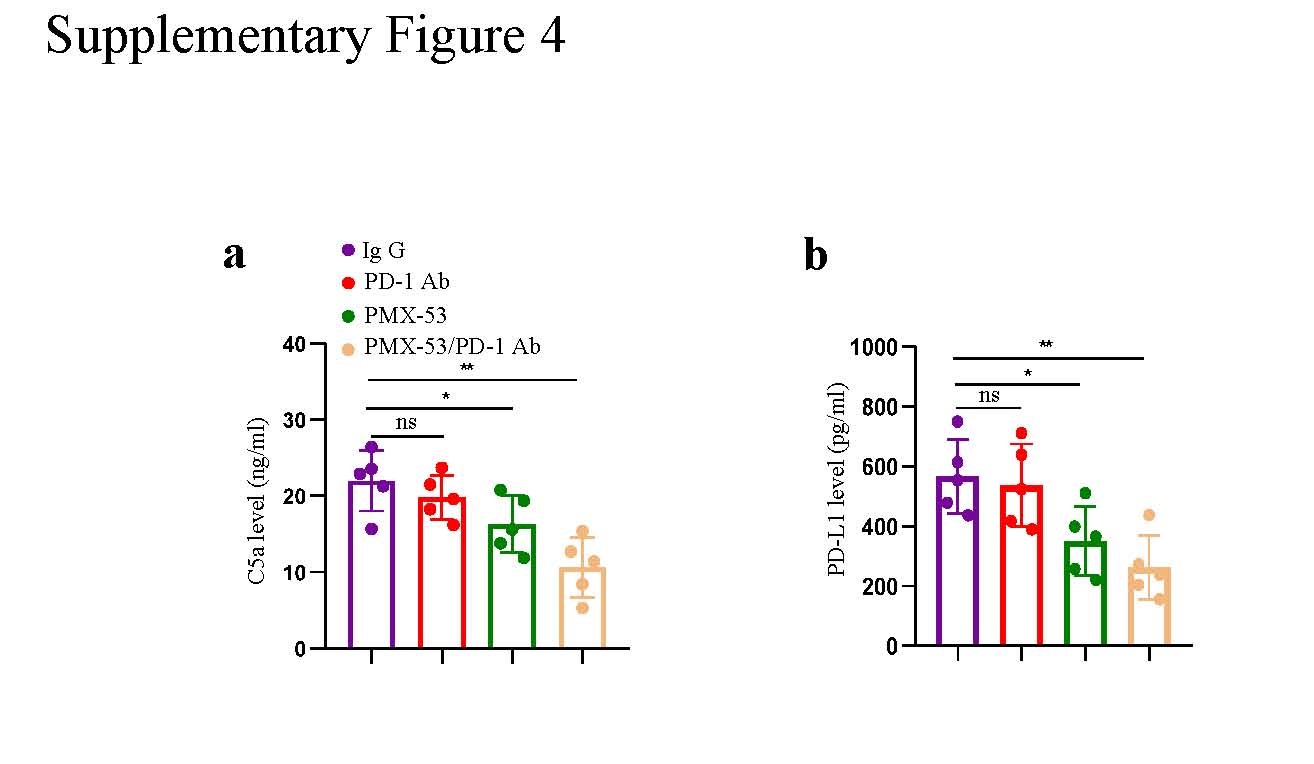
Considering the promising effects of PMX-53 in inhibiting the PD-L1 expression of macrophages and reversing CD8+ T cell dysfunction, we speculated that the combination therapy of PMX-53 and anti-PD-1 antibody could inhibit the progression of NSCLC driven by circASCC3 and improve the efficacy of anti-PD-1 therapy. We performed an in vivo investigation of the anti-PD-1 antibody and PMX-53 combination therapy in an LLC-circASCC3 xenograft subcutaneous model in C57BL/6 mice. The anti-PD-1 antibody was administered on days 1, 4, 7, 10, and 13, and PMX-53 was administered daily when the tumor size reached 100 mm3 (Fig. 6a). The in vivo results showed that the combination therapy significantly inhibited tumor growth compared to PD-1 or PMX-53 monotherapy (Fig. 6b, c). In addition, the combination therapy also increased survival and reduced lung metastasis compared to either single agent (Fig. 6d, e). The C5a and PD-L1 level in the plasma of mice were significantly reduced after the combination therapy of anti-PD-1 monoclonal antibody and PMX-53 (Supplementary Fig. 4a, b).
Moreover, flow cytometry analysis showed a significant reduction in M2-type macrophage infiltration and the expression of PD-L1 in macrophages in the tumors of mice treated with anti-PD-1 monoclonal antibody and PMX-53 compared to the respective levels in the tumors of mice treated with monotherapy (Fig. 6f, g). In addition, the tumor-infiltrating CD8+ T cells were apparently increased, and the cytotoxic function of the CD8+ T cells was also obviously improved, as reflected by the increased infiltration of IFN-γ+CD8+ T and GZMB+CD8+ T cells in tumors after the combination therapy of anti-PD-1 antibody and PMX-53 (Fig. 6h). Thus, the in vivo results in an immunocompetent mouse model suggest that the abnormal expression of circASCC3 is involved in the immune evasion of NSCLC cells and that the C5aR1 inhibitor PMX-53 can improve the efficacy of anti-PD-1 immunotherapy.
Expression of circASCC3 correlates with the efficacy of anti-PD-1 therapy in NSCLC patients
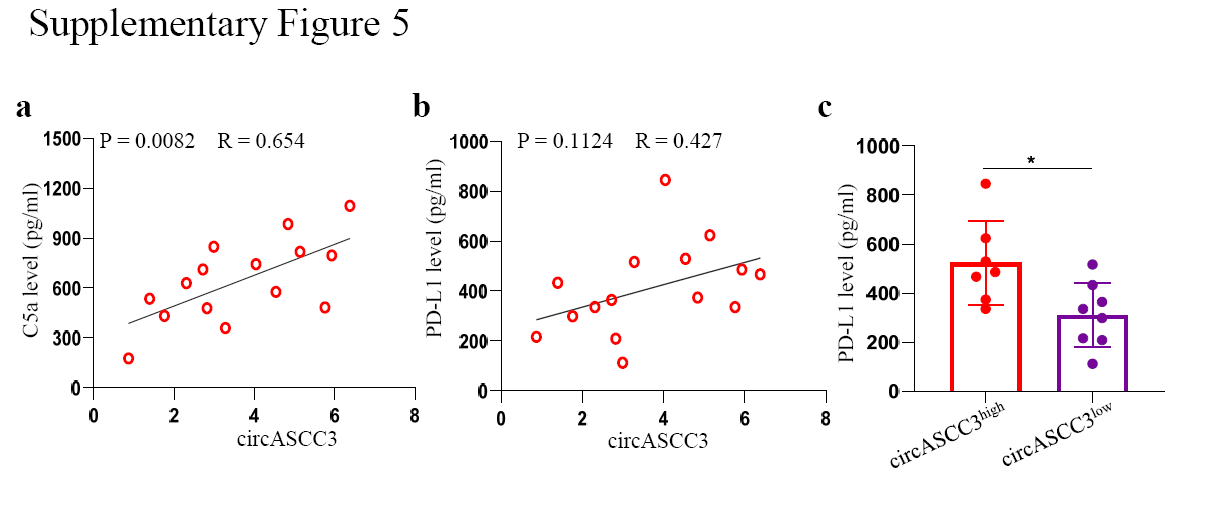
To further validate our results in NSCLC patients, we retrospectively analyzed the data from 15 NSCLC patients in clinical stage III or IV according to the 8th TNM staging system who received anti-PD-1 immunotherapy (Opdivo). Following the RECIST1.1 evaluation, 7 patients had progressive disease (PD), 6 patients achieved stable disease (SD), and 2 patients were classified as having partial remission (PR). Next, the expression of circASCC3 was evaluated in biopsied tissues using qRT-PCR. The results showed higher expression of circASCC3 in patients with PD than in patients with SD or PR (Fig. 7a). Among the patients with high circASCC3 expression, only 2 patients had SD, and 5 patients were classified as having PD. In patients with low circASCC3 expression, 2 patients had PD, 4 patients had SD, and 2 patients were classified as having PR (Fig. 7b). In addition, the expression of circASCC3 was positively correlated with the expression of C5 (Fig. 7c and Supplementary Fig. 5a). While no relationship was found between the PD-L1 in the plasma and the expression of circASCC3, the concentration of PD-L1 was higher in patients with high circASCC3 expression (Supplementary Fig. 5b, c). The mIF results of tumor tissues showed lower infiltration of CD8+ T cells but higher infiltration of CD163+PD-L1+ macrophages in tumors with high expression of circASCC3 (Fig. 7d).
Moreover, the imaging results also showed a similar pattern. The results of contrast CT imaging of two representative patients are shown in Fig. 7e. In the high circASCC3 group, 5 patients showed tumor growth, and only 2 patients had slight tumor shrinkage; in contrast, 6 patients in the low circASCC3 group had tumor shrinkage, and only 2 patients had tumor progression after treatment (Fig. 7f). In addition, a negative correlation was presented between the expression of circASCC3 and shrinkage of tumor size (Supplementary Table 1). Next, we further investigated the relationship between circASCC3 and the prognosis of 15 patients. PFS and OS were prolonged in the low circASCC3 group (Fig. 7g). Taken together, these preclinical results reveal an association between circASCC3 and immune resistance and provide a potential strategy for overcoming resistance to anti-PD-1 immunotherapy in patients with NSCLC.
Figure 8 shows a schematic summarizing the results of this study. In NSCLC, highly expressed circASCC3 can sponge miR-432-5p, which further upregulates the expression of downstream C5a and induces EMT to promote the progression of NSCLC. On the other hand, the activation of the C5a/C5aR axis in macrophages in the TME induces macrophages to transition toward an immunosuppressive phenotype and elevates the expression of PD-L1, which further leads to the dysfunction of CD8+ T cells.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}