In this study, we determined the polarization of macrophages using six surface polarization markers, including CD40, CD80, CD16, CD163, CD206, and CD200R. CD40 and CD80 were considered biomarkers for M1 macrophages as they are highly expressed in M1 macrophages but not in M2 macrophages [28]. CD16 and CD163 were regarded as IL10 M2 biomarkers [29], while CD206 and CD200R were have been wildly detected as IL4 M2 biomarkers [33]. Compared to cells from healthy donors, cells from rheumatoid arthritis patients expressed a higher level of CD40 and CD80, yet, they had a lower level of CD16, CD163, CD206, and CD200R. Our study demonstrated that compared to macrophages from healthy donors, macrophages from rheumatoid arthritis patients have a higher level of M1 polarization, but had a lower level of M2 polarization. We suggested the higher level of pro-inflammatory M1 polarization of cells is associated with the inflammatory in rheumatoid arthritis patients, on the other hand, the lower level of anti-inflammatory M2 biomarkers in rheumatoid arthritis patient cells indicated that the patient cells had a lower inflammatory.
We also explored the role of the CDKN2B-AS1/MIR497/TXNIP axis in rheumatoid macrophages. Surprisingly, in macrophages from healthy donors, there was no correlation among CDKN2B-AS1, MIR497, and TXNIP mRNA levels. These results suggested that CDKN2B-AS1/MIR497/TXNIP axis might not play role in healthy donors. However, in macrophages from rheumatoid arthritis patients, CDKN2B-AS1 was significantly negatively correlated to MIR497, CDKN2B-AS1 was significantly positively correlated to TXNIP, and MIR497 was significantly negatively correlated to TXNIP. These results indicated that CDKN2B-AS1/MIR497/TXNIP axis was activated only in rheumatoid macrophages but not in macrophages from healthy donors. We suggested that when the CDKN2B-AS1, MIR497, and TXNIP at a normal level, they are not able to significantly interacted with each other. Only when they are at an abnormal level can they regulate each other. This might also involve some unidentified feedback mechanisms that can be explored in the future.
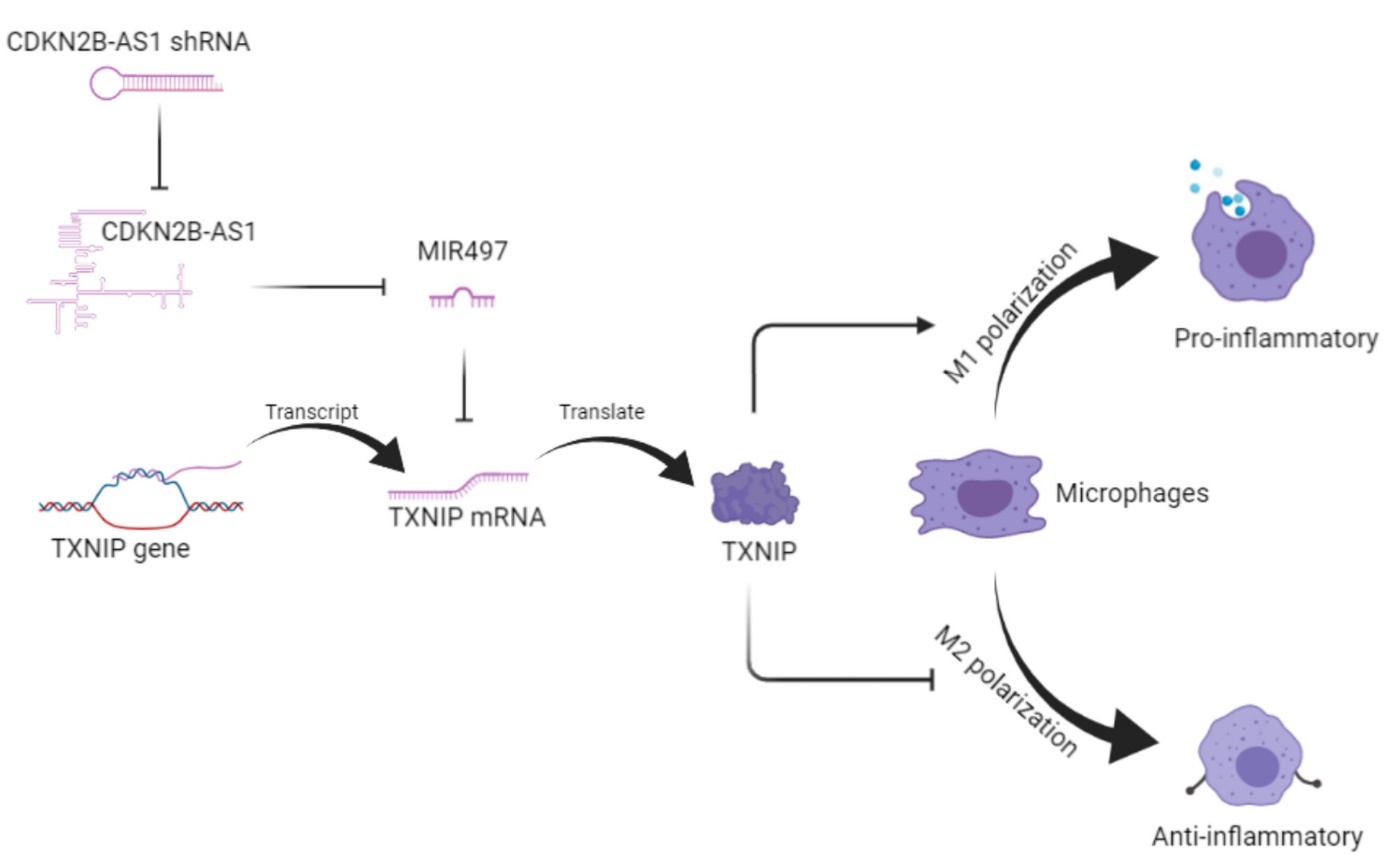
To validate the regulation of the CDKN2B-AS1/MIR497/TXNIP axis in the polarization of macrophages, we interfered with the expression and function of the axis from upstream to downstream. The role of the CDKN2B-AS1/MIR497/TXNIP axis in macrophages was summarized in Fig. 7A. Our study demonstrated that CDKN2B-AS1 regulated TXNIP through MIR497. The complementary binding sites of the CDKN2B-AS1/MIR497/TXNIP axis were shown in Fig. 7B. Notably, because the complementary binding sites only have seven complementary base pairs, the CDKN2B-AS1/MIR497/TXNIP axis might only have relatively weak interaction, which might account for the deregulation of this axis in health donor cells. The sponging of MIR497 by CDKN2B-AS1 can inhibit the level of endogenous MIR497. Most of the miRNAs play roles in cells by their inhibition toward their target mRNA [17]. In this study, as MIR497 can directly suppress the translation of TXNIP mRNA to TXNIP protein, the CDKN2B-AS1/MIR497/TXNIP axis regulated the level of TXNIP. Our study also revealed that TXNIP had a novel regulation role in the polarization of macrophages. TXNIP can facilitate M1 pro-inflammatory polarization but inhibited M2 anti-inflammatory polarization in macrophages, thereby positively regulated the inflammation in rheumatoid arthritis.
However, the monocytes in this study were in vitro differentiated, which were different from the macrophages obtained from patients that might be affected by the internal environment of the patient. Yet, this study is focusing on the gene expression and the roles of the CDKN2B-AS1/MIR497/TXNIP axis. We think the effect of the genotype of the cells was more critical than the internal environment that induced the cell differentiation. We presumed that the cells from patients have a genotype that results in the onset of rheumatoid arthritis and the genotype would not be affected by the internal environment of the patient. Thus, we only studied in vitro differentiated cells. In addition, the mechanism of TXNIP regulating the polarization of macrophages has not been studied in this study. One potential mechanism involves might be the ion channel regulation in macrophages. Ion channels have been found to be critical in many cells [34–37], some macrophages expressed ion channels for their functions [38]. TXNIP has been found to be involved in some ion channel regulations [39]. Regarding the clinical treatment of rheumatoid arthritis, many anesthetics have been applied in therapy to relieve rheumatoid arthritis pain [40, 41]. These drugs usually have multiple ion channel targets [42, 43]. Many anesthetics can also have multiple ion channel targets[42] and regulate the immune system [44, 45] that might interfere with rheumatoid arthritis. Whether the use of these anesthetics interacts with macrophages in rheumatoid arthritis is not clear. In addition, the potential role of the CDKN2B-AS1/MIR497/TXNIP axis in other bone diseases is also interesting. Studies have found that TXNIP is closely associated with bone metabolism [46–48]. Rheumatoid arthritis and osteoporosis have been two of the most pervasive bone diseases [49, 50]. For example, many pathways have been proposed to regulated osteoporosis [51–54], but whether osteoporosis can be regulated by the CDKN2B-AS1/MIR497/TXNIP axis can also be explored in the future.
{kind=link}