A comprehensive profile of structural and functional dynamics of fungal community during fruit waste composting was investigated. For this purpose, fruit waste was composted in a windrow setup. Representative samples were collected at varied range of incubation temperatures during composting period and employed to physicochemical and microbiological culture dependent and independent analysis. Time-series data revealed that variation in fungal load is temperature dependent that influenced morphotypes’ shifts during different stages of composting. Shifts in abiotic factors, availability of accessible nutrients, water loss, pH and electrical conductivity participated in the transition of community and compost maturity. Culture-based analysis showed rich microbial compost community, dominant with Aspergillus, Mucor, Rhizopus and Penicillium. Denaturing gradient gel electrophoresis analyses demonstrated the prevalence of diverse community in compost with detectable bands corresponding to Penicillium at mesophilic temperature while undetectable bands corresponding for Aspergillus. Succession in microbial community was observed during composting as with temperature variations. Illumina Miseq revealed fungal diversity including Mortierella sp from phylum Zygomycota as the most dominant fungi and Coprinopsis sp as second dominant from Basidiomycota, mainly associated with lignocellulosic degradation. Moreover, Aspergillus fumigatus (ADIF1) was found as the most promising cellulase and pectinase producers at higher temperature showing its potential for efficient environmental management utilization. Current findings suggest that transformation of fruit waste into seed germination friendly compost that can be used as an efficient organic fertilizer and incorporation of sensitive molecular technique suggests the transition of microbial community and improvement in microbial diversity.
Research Article
Succession and Catabolic Properties of Fungal Community During Composting of Fruit Waste at Sub-Tropical Environment
https://doi.org/10.21203/rs.3.rs-704507/v1
This work is licensed under a CC BY 4.0 License
Journal Publication
published 10 Jan, 2022
Read the published version in Waste and Biomass Valorization →
You are reading this latest preprint version
Even though numerous studies have looked at the significance of microbial communities in composting, there is still potential for more research into microbial populations due to their variability based on feedstock and environmental conditions. Therefore, this study is envisaged to determine the fungal diversity of organic compost using local indigenous waste via standard plate count, DGGE and next generation sequencing. It was observed that using a culture-based approach alone is insufficient, and that combining it with sequencing uncovered an uncultured fungal population that is catabolically diverse and plays a prominent role in producing germination friendly compost. We believe that the manuscript findings will provide valuable insights into the microbial-driven composting of fruit waste, that can aid in rapid co-composting of lignocellulosic waste.
Management of biowaste is a huge problem in low- and middle-income countries. These countries generate around 54% of biowaste of their total waste [1]. Open dumping of these wastes in landfill sites results in air and ground water pollution through leaching which poses a high risk to human health. Composting is a cost effective and easy method to manage waste in developing countries. It not only reduces the waste volume but also provides organic fertilizer that could be used to enhance soil fertility [2]. The composting process is a combined activity of bacteria, fungi and actinomycetes. Usually bacterial community outnumbers the fungal community during mesophilic and thermophilic phase of composting [3–5], owing to their quick generation time. The other reason that favors the bacterial growth is physicochemical properties of feedstock. For example, the presence of Lactobacillus in initial stage indicates influence of low pH on their growth [6]. Similarly, low pH also favors the growth of yeasts in early phase of composting [7]. But strikingly, fungi being the efficient degraders of recalcitrant substance in the later stages of composting have not been thoroughly studied earlier and its importance has remained neglected. Recently, several studies attempted to study fungal succession by incorporating molecular methods like denaturing gradient gel electrophoresis (DGGE) and pyrosequencing [7,8]. These studies revealed that microbial succession is depends upon the feedstock and the method of composting used to manage waste. The existence of standard microbiota during composting is not possible since it’s a diverse process which is highly dependent upon feedstock used and technique of composting applied and does not operate in standard environmental conditions but the variation in physical and biological factors fairly follows the same trend [9]. It has been stated that different composting methods, waste sources, and environmental parameters result in different microbial communities, which is why it is critical to isolate and study their participation to generate high-quality compost [7,10–13]. Considering these facts, in this study we planned to use fruit waste as a main feedstock for composting. Fruit waste contains high content of Nitrogen and other macronutrients like Phosphorous and Potassium which upon degradation could play role in enrichment of nutrient deficit soils. These macronutrients play a major role in plant growth. Addition of micronutrient rich compost in to soil increases the abundance of saprophytic fungi [10].
The enrichment of the soil can be done through composting of these organic waste such as fruit peels, soybean residues and food waste since they are easily available and contain high nutrients (K, P, N), but most of the time these wastes are not utilized effectively but are disposed of through burning. To the best of our knowledge, no study in Southern Pakistan was done to provide a comprehensive idea of fruit waste composting and the microorganisms involve during overall process of degradation. Since local environmental conditions play a very important role with the feedstock and physical parameters used, we were aiming to observe the variation in microbial community during composting under such conditions.
In this study, microbiological and analytical methods were used to access the biodiversity of microbial communities. To achieve this aim, following objectives were targeted, a) to assess the rate of biodegradation and time required to compost maturation; b) to monitor environmental parameters and study its transitional effect on microbial diversity; c) to study structural and functional composition of resident microbial communities and their dominancy of isolates over various phases of composting and d) to assess phytotoxicity of mature compost.
4.1 Composting Setup and Physical Parameters:
The study followed a windrow setup for the degradation of fruit waste through composting. Fruit waste was used as a key substrate along-with green leaves, fresh grass clippings and untreated sawdust to achieve a C:N ratio of 25-30:1. C:N ratio was achieved through a volume-based ratio calculation of each substrate. These substrates were also amended with garden soil that acts as starter and aid in composting. Prior to composting, each layer of substrate was moistened enough to adjust the moisture content to 60-65 %. This setup was done on an unpaved ground in green house conditions at the backyard of the department of Microbiology, University of Karachi (24°56′ 24′ N 67°7′ 6′ E) for a period of five months, i.e., March to August 2017 and exposed to environmental temperature ranging from 29-36 ºC. This pyramid shape windrow pile (105cm long x 85cm wide x 110cm tall) was aerated and moistened periodically based on its temperature and moisture profile which was monitored through mercury thermometer and oven drying method, respectively. Temperature and moisture content was monitored from different sections within a pile; surface (10cm from top) and center (55cm from top) of pile.
4.2 Sampling Strategy and Chemical Analysis:
Efficient compost has certain established physicochemical conditions. Samples from different positions of compost pile were collected to make a composite representative sample of each sampling day and then it was subjected to physicochemical and fungal community analysis. All the analyses were performed in triplicates. Total carbon (TC) was determined gravimetrically after ignition in oven at 750 °C for 2 hours and Total Nitrogen (TN) was estimated using Kjeldahl method [11].Carbon to Nitrogen (C:N) ratio was calculated as the quotient obtained after dividing Carbon content to Nitrogen content [12]. The pH and Electrical Conductivity (EC) were measured using pH and conductivity meter (Hach, USA). Macronutrients (Calcium, Magnesium and Phosphorus) were quantified through Atomic Absorption Spectrophotometer using an acid-treated compost samples [13].
4.3 Culture-dependent Fungal Community Analysis:
A suspension of compost using 10g of sample and 100ml of Phosphate buffer saline (PBS) was prepared and shaken for 30 minutes to homogenize the sample. The viable fungal fraction of each sample was enumerated using ten-fold serial dilution and 100µl of serial dilutions (10-1 to 10-4) were inoculated on Sabourauds Dextrose Agar (SDA, sigma UK) medium. Inoculated plates were incubated in triplicates at five different temperatures (25 °C, 37 °C, 45 °C, 50 °C and 55 °C) to have complete mesophilic and thermophilic profile of fungal community. In case of mesophilic fungal cultivation (25 °C, 37 °C), the incubation time was 5-7 days till no increase in number of fungal cultures was observed. For thermophilic fungi (45 °C, 50 °C and 55 °C) the incubation time was 3-5 days. The viable fungal population was expressed in CFUg-1 of compost sample. The fungal colonies that differ in macroscopic characteristics were sub-cultured on SDA and were used for morphological grouping [14] to estimate their diversity and then followed with microscopic observations to confirm the isolates. The fungal isolates belong to different genera were again sub-cultured and purified for molecular identification based on its sequencing. Prior to DNA extraction, these distinct fungal colonies were treated with liquid Nitrogen and then genomic DNA extraction was carried out as described elsewhere [15]. Amplification of ITS1-5.8S-ITS2 region of fungal rRNA gene was performed using primers ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) and ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) [16]. The reaction proceeded for 35 cycles with denaturation at 95 °C for 30 s, annealing at 55 °C for 60 s and elongation at 72 °C for 120 s. Amplified PCR products were purified using QIAquick PCR Purification Kit following manufacturer’s instructions (Qiagen, UK). Sanger dideoxy sequencing of purified amplicons were carried out via Macrogen. Accession numbers were obtained from National Center of Biotechnology Information database.
4.4 Culture-independent Fungal Community Analysis via DGGE
For Fungal community analysis by DGGE, genomic DNA was extracted using DNeasy Powersoil Kit (Qiagen, UK) according to manufacturer’s Instructions, followed with amplification of ITS1 region using primers GM2 (5′CTGCGTTCTTCATCGAT-3′) and JB206 (5′CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGGAAGTAAAAGTCGTAACAAGG-3′) [17]. PCR products were confirmed through a 1.5 % (w/v) agarose gel with a Hyperladder I (Bioline, UK) at 100 V for 1 h electrophoresis. Amplicons from three replicates were pooled and purified using a Qiagen purification kit according to the manufacturer’s instructions. DGGE was performed according to the protocol described elsewhere [18]. The position and intensity of the bands was quantified using Phoretix ID Advanced V5.00 software (Nonlinear dynamics, UK). In order to standardize and allow comparisons to be made between different gels, pooled PCR products amplified from the DNA of Penicillium chrysogenum, Rhizopus oryzae, Aspergillus niger, Aspergillus fumigatus and Aspergillus flavus were included as a species ladder on every gel.
4.5 Culture-independent Fungal Community Analysis via Illumina Miseq
Microbial community DNA was extracted from 1g mature compost (week 20) using DNeasy Powersoil kit (QAIGEN) according to manufacturer’s instructions with modifications. Vortexing time was increased for proper lysis step i.e., 20 minutes and centrifugation was done at 12,000 rpm for 2 minutes. Eluted DNA was visualized using UV transilluminator (UVP's patented Benchtop Transilluminator). Gel Electrophoresis was performed in 1% agarose gel stained with 1.5μL ethidium bromide. Wells were loaded with 3μL of DNA sample. Gel was run for about 15 minutes along with DNA ladder at 100 volts. Samples were stored at -20ºC till required. Extracted DNA samples were sent to BGI, Hong Kong for library construction, Internal Transcribed Spacer (ITS) amplicon sequencing and bioinformatics analysis. Fungal ITS 1 region sequencing was done by using illumina MiSeq system that generates 300 paired ends (50,000 TAGS).
4.6 Catabolic Profiling:
Selected fungal isolates were also screened for their cellulase and pectinase activity at mesophilic (30 ºC) and thermophilic (50 ºC) temperatures. Well diffusion method and agar plate method were employed to determine cellulolytic and pectinolytic activities, respectively. In well diffusion method, fungal cultures were grown on carboxymethyl cellulose (CMC) broth [19] at 150 rpm in a shaking incubator for 72 h and harvested by filtration through Whatman no. 1 filter paper that resulted in cell free culture supernatant. Test media (cellulose 1 %, Congo red 0.5 %, agar 1 %) was bored and 40µl of cell free culture supernatant was added and incubated for 24 h. Plates were stained with Congo red dye (1%) for 15 min followed by de-staining with 2X NaCl for 15 min [20]. The halo around well is indicative of cellulolytic activity. For quantitative analysis of cellulase, cell free culture supernatant was used as a crude enzyme to perform Endoglucanase assay [19]. Crude enzyme (50 µl) was added to equal volume of 1 % (w/v) CMC in 50 mM sodium citrate buffer, pH 4.8. The tubes were incubated for 30 min at respective temperatures (30 ºC and 50 ºC) followed by the determination of reducing sugar. One unit of Endoglucanase activity was defined as the amount of enzyme that can hydrolyze CMC and release one micromole of glucose under standard assay conditions [19]. In Agar plate assay, isolated fungal colony was spotted through needle on media plate containing Minimal Salt Medium with 1 % Pectin [21] and left for overnight incubation. Pectinolytic fungi were identified through flooding the incubated plates with Iodine crystals resulting in the production of clear halo around colonies.
4.7 Phytotoxicity assessment:
The maturity of compost was assessed by carrying out phytotoxicity assay as described by Haq et al [13] and Ansari et al [22] using green mung bean (Vigna radiata) as test seeds. For this purpose, seed germination was performed in a petri dish lined with sterilized cotton and filled with aqueous extract of compost (5 ml). This compost extract was obtained by mixing 10g of compost sample with 100ml of distilled water, shaking it for 30 mins at room temperature to homogenize it and then filtering it through Whatman no. 1 filter paper. In control plates, autoclaved distilled water (5 ml) was used. Then six mung beans were placed in each plate and incubated in dark for five days. After incubation time, germination index (GI) was calculated by the root length and the percentage of germination of selected test plant seeds compared to a control using following formulae.

4.8 Statistical Analysis:
Descriptive statistics (Mean and standard deviation) were calculated for triplicates of each sample by using SPSS software (IBM SPSS STATISTICS 20). The observed data were also subjected to one-way analysis of variance (ANOVA) to determine the differences of physicochemical and microbiological analysis in each sample during composting. Significant acceptance was considered at P <0.05. Principal Component Analysis (PCA) of bands on DGGE gel was performed using Multi-variate Statistical Package (MVSP) version 3.22 (copyright © 1985-2003 Kovach computing services).
5.1 Changes in Physicochemical parameters:
Temperature of compost pile (Surface and Centre) and environment was monitored throughout the composting process that started on zero day and ended on 146th day. Changes in temperature profile were observed in Figure 1, depicted that as compared to the ambient temperature. 28 °C, the temperatures of center and surface of pile was started to increase overnight and peaked to 42 °C (Surface) and 49 °C (Center) on 7th day indicating thermophilic phase. This rise in temperature was due to quick decomposition of easily-biodegradable organic compounds [3]. With progressive degradation, temperature of the compost pile gradually dropped and thereafter remained consistent with time to ambient temperature. As shown in Figure 2, the process was started from 67 % of water content and ended at significant loss to 22.4 % (surface) and 31.5 % (center) by the end of composting. The difference in moisture loss between surface and center of pile was due to more exposure of surface heap to sun that led to evaporation [23], while water in the center remain trapped.
The variation in pH and EC throughout the composting process is shown in Figure 3. In the case of pH, the feedstock was slightly acidic initially, i.e., pH 6.0 due to acidic nature of citrus fruits waste. As acidic feedstock underwent degradation, rise in pH was observed. This significant rise (p<0.05) to alkaline pH 8.1 was observed on 7th day, possibly due to protein hydrolysis or ammonia formation. From 7th day onwards, pH declined to neutral (7.0) and then stabilized at pH 6.0 till curing. In the case of EC, at the time of piling, the compost conductivity was 0.16 mScm-1 and with the progressive degradation it ended at 0.26 mScm-1 with a significant increase (P<0.05).
The changes in C:N ratio, and macronutrients that includes calcium, magnesium and phosphorus are tabulated in Table 1.Composting was started with a C:N ratio of 31:1 which is an optimum range for composting. Over the composting period, C:N ratio declined consistently and significantly (P<0.05) and ended at 19:1. Variations in macronutrients content was evident as in the case of Calcium and Magnesium a significant increase (P<0.05) was observed in samples collected from initial and final stages of composting. Whereas Phosphorus content was found consistent throughout the process of composting.
The efficiency of cured compost can be assured by the physicochemical parameters. The overall results suggest that thermophilic range was obtained just in the first week of degradation, the same time pH rose as well. The first seven days also showed significant rise in N content with a drop in C content and C:N. The same period showed sharp rise in P and other macronutrients. Decrease in organic matter suggests the degradation of waste material in compost that led to the release of nutrients. The heat generation during active phase of composting and presence of optimum concentration of nutrients in matured compost indicated the efficiency of compost.
5.2 Changes in Fungal Community during Composting via Culture-based Technique:
The count of viable fungi at different time points during composting showed an increasing trend at mesophilic temperatures (Figure 4, P<0.05). In the beginning, fungal count at 25 ºC was 5x106 ( + 1 x105) CFUg-1that significantly increased (P<0.05) to 3.76 x107 ( + 2.5 x106) CFUg-1in second week of composting and it ended at 1.5 x107 ( + 1.7 x107) CFUg-1 with several fluctuations in count. Similarly, at 37 ºC, initial count was 2.0 x106 ( + 2.5 x105) CFUg-1 that substantially increased to 4 x107( + 1 x106) with time (day 15, P<0.05). In contrast, thermophilic temperatures (45 °C, 50 °C and 55 °C) showed variations in their trend with the rise in temperature. At 45 °C, the fungal count was 1.46 x106 ( + 3.05 x105) CFUg-1 that significantly increased during thermophilic phase and reached to 5.3 x107 ( + 3.05 x106) CFUg-1 and ended at 7.6 x107 ( + 5.7 x106) CFUg-1 conferring an overall increasing trend. However, at 50 °C and 55 °C, the fungal count had significantly declined from 2.13 x106 ( +1.5 x105) to 1.4 x105 ( + 3.0 x104) CFUg-1 and from 1.8 x106 ( + 8.1 x105) to 7.0 x104 ( + 1.0 x103) CFUg-1 respectively, with the increase of pile temperature in the first week of composting. It was found that variation in fungal viable load existed throughout the composting period of 146 days. This variation was temperature- related which means the fluctuation in fungal count was not only observed within the temperature at different time points of composting process but also among incubated temperatures.
5.3 Changes in Fungal Communities during composting via culture dependent technique:
Fruit waste was primarily degraded by filamentous fungi, which could be grouped based on colonial characteristics. Therefore, relative progression of each morphotype was calculated by counting the distinctive colony in each sample at various time intervals and at various incubation temperatures. The dynamics of fungal communities at all incubation temperatures are demonstrated in Figure 5. The findings show that Yeasts predominate in the early stages of composting at mesophilic temperatures due to presence of low pH and high moisture content. Whereas, on the later stages at mesophilic temperatures (25°C and 37°C) the phenotypic diversity of the fungal population was observed. At 25 °C, yellow powdery colonies with a white periphery and some black spores predominated, as did white cottony small-sized colonies. Similarly, at 37°C, white mycelial medium-sized colonies that matured to a greyish color, extremely dense black sporulating colonies, and gray powdery colonies were found to be similarly dominant morphotypes. However, no such diversity among fungal morphotypes was observed at thermophilic temperatures (45°C, 50°C, and 55°C). At 45°C, 50°C, and 55°C, the major morphotypes were green centered colonies with white periphery, white cottony colonies that turned brown over time, and white mycelial cottony colonies that swarmed over the entire growth medium on plate.
The results proposed that variation in colonial characteristics of fungal isolates were dependent on the temperature and the local abiotic conditions of the compost.
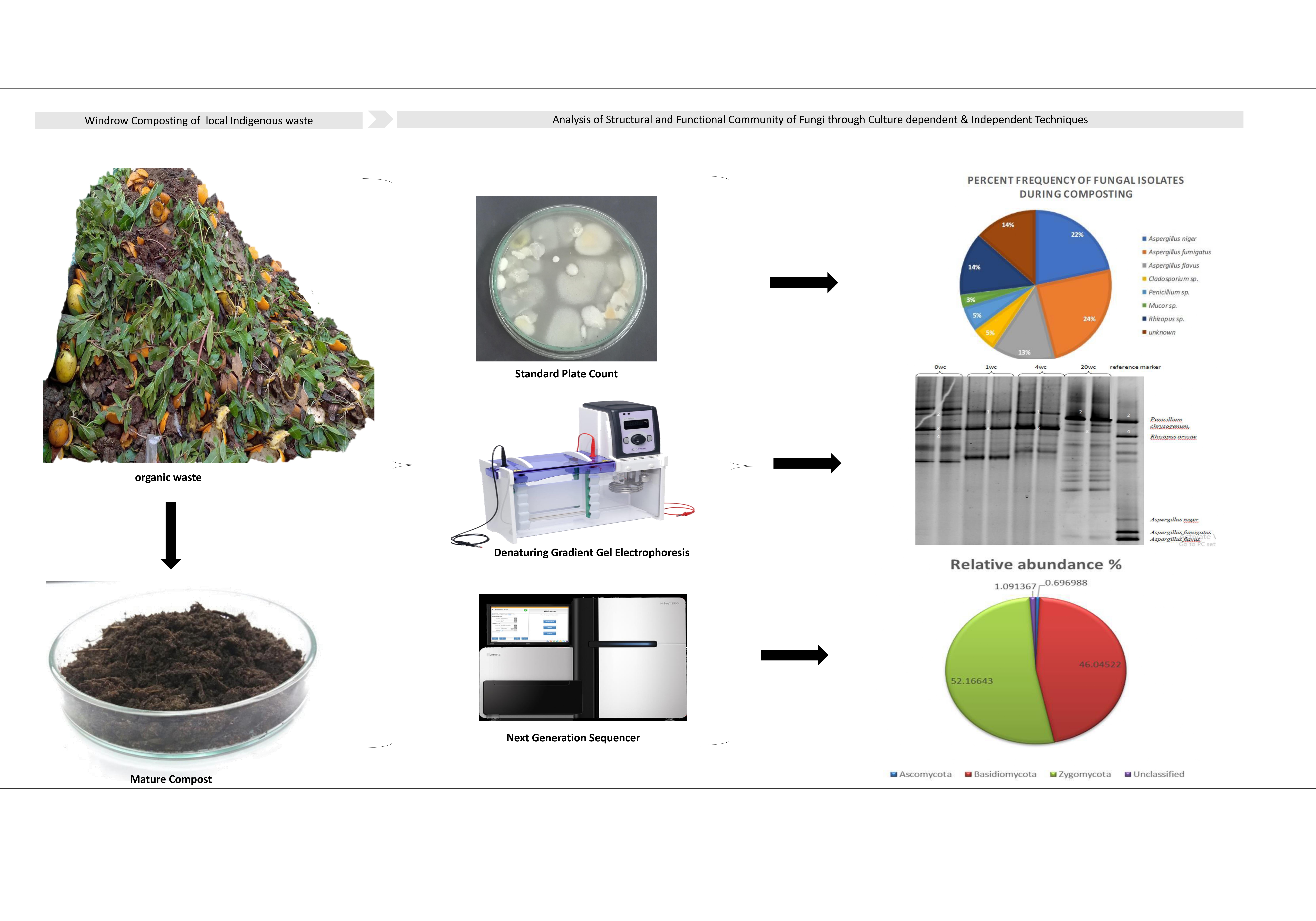
These morphotypes were then analyzed under a microscope and classified into genera. With reference to Zafar et al. [26], this grouping and probable identification of isolates was done based on culture and microscopic morphology. Among those genera, Aspergillus spp. have dominated for the entire 146-day composting period (Figure 6). This demonstrated the fungal community's active participation in fruit waste degradation through composting.
5.4 Changes in Fungal Community during composting via culture independent technique:
DGGE revealed that in the compost, the fungal population diversity was moderate initially when the pile was first formed. After week 4 and week 8, the community profile was very low that could be due to higher temperature, where bacteria play a major role in the degradation of organic waste (Figure 7). As the temperature goes down and compost gets mature, the diversity of fungal community increases. PCA analysis of Boolean data (presence/absence of bands on DGGE, 0 or 1), confirms the succession of fungal community dynamics from week 0 to week 20. The PCA analysis clearly demarcates the three samples into thermophilic, mesophilic, and curing phases of compost (Figure 8 & 9). Three bands were recovered from first two stages (week 4 and week 8) at different positions and thirty-three bands were recovered in the last sample suggesting the richness of community. Band 1 only appeared in the week 4 and week 8 when temperature of the compost surface was around 40-50 °C. Band 2 corresponding to Penicillium chrysogenum only appeared in samples at mesophilic temperature range i.e., week 0 and week 20. Band 3 was constantly present throughout the composting process with thickness variability, which could be attributed to its abundance. Interestingly, the fungal genus corresponding to this band was either not or hardly recovered through culture-based technique implied. Band4 corresponds to Rhizopus oryzae which was frequently recovered in culture-based technique but appeared in low abundance through DGGE (Figure 7).
Another interesting observation revealed through DGGE is that the species recovered regularly through culture-based technique were not prominent in the molecular analysis. This could be because of their high germination rate exploited during culturing or biasness of ITS primers during community PCR. DGGE also revealed that there are many other species present that were not recovered through culturing.
5.5 Biodiversity of fungal communities associated with UTBC by Illumina Miseq:
A total of 131,853 sequences were obtained after removing all poor quality and short reads data and were clustered into 47 different OTUs at 97% sequence similarity. In order to know the complexity of sample, alpha diversity was applied through indices such as observed species (Figure 2 supplementary file) Chao1, Shannon- index and Simpson (Table-2 supplementary file). Complexity of sample is proportional to all indices except that of Simpson value. Species richness of community is usually reflected by observed species and Chao1 value. Shannon index together with Simpson value provide an idea about Species diversity in community that ultimately depends on species richness and evenness. The rarefaction curve indicated the produced data was enough to cover all 47 species in fungal community. The straightening of curve suggested, data produced was enough to study diversity of sample.
Species annotation showed that sequences were associated with 3 phyla including Zygomycota, Basidiomycota and Ascomycota and among phyla, Zygomycota was found as the most dominant one with relative abundance 52.16 % (Figure 10). On species level, Mortierella sp. and Rhizomucor sp. were identified. Basidiomycota revealed as the second dominant phyla with relative abundance 46.04 %, identified species were Coprinopsis sp., Coprinellus canistri & Coprinus cordisporus. Results indicated less abundance (0.69 %) of Ascomycota detected with Scutellinia sp., Schizothecium carpinicola, Penicillium sp., Preussiaterricola, Arthrobotrys thaumasia, Zopfiella sp., Montagnulaceae sp., Trichoderma longibrachiatum, Aspergillus sp., Dactylella sp., Monascus purpureus, Myrmecridium schulzeri, Emericella sp. and Fusarium sp. phylogenetic tree at Genus level from different phyla is shown in Figure 11. The identified illumina sequences were submitted to NCBI GenBank for accession numbers as mentioned in (Table 3 Supplementary file)
5.6 Catabolic Profiling of fungi:
Since, the compost pile has been made from fruit waste, leaves, and saw dust, it became a good source of pectin and cellulose, and therefore fungal strains were investigated for pectinolytic and cellulolytic activities.
Carboxymethyl cellulose was used as carbon source for screening of cellulase producers. Cellulolytic activity of all the fungal isolates was determined by its ability to degrade β-D-glucan in medium. At mesophilic temperature, fungal isolates that possessed cellulolytic activity were Aspergillus flavus, Aspergillus fumigatus, Aspergillus niger, Penicillium sp. and Cladosporium sp. Whereas, A. fumigatus (ADIF1, ADIB8 and ADIK2) grown and exhibited cellulolytic activity at thermophilic temperature. The highest cellulolytic activity was shown by ADIF1 (A. fumigatus) at 50 ºC, i.e., 4.40 IUml-1, via Endoglucanase assay. The cellulolytic activity of fungal strains was much higher at thermophilic temperature compared to mesophilic temperature. To our understanding, high temperature was optimum for the growth and secretion of cellulases by fungal strains.
Pectinase production was observed as halo around fungal colony due to depolymerization of pectin in growth media by exposing with iodine crystals. In this study, A. flavus and Penicillium sp. were observed as pectinase producers at 30 ºC and A. fumigatus exhibited pectinolytic activity at 50 ºC.
The results presented in Table 2 indicated the importance of genus Aspergillus for their hydrolytic activity and their detailed study could result in a development of better strains that can effectively degrade complex substances and has diverse industrial applications.
All the representative fungal isolates that were selected and subjected to enzymatic analysis were chosen for identification through amplification of ITS gene (Table 1 supplementary file). DNA of Cladosporium sp. could not be recovered that is why its molecular identification was not performed. Overall, the results suggest that temperature variation in the incubation and composting itself favors the growth of certain species at specific time intervals of composting process. Investigation at five different temperatures, helped the comprehensive exploration of microbiological niche found in compost.
5.7 Phytotoxicity of compost:
Germination Index (GI) was calculated by combining the measure of relative seed germination and relative root elongation. From the results obtained, GI of compost was 83.3 % hence indicating its maturity and absence of phytotoxic compounds (Table 3), which is higher chemical fertilizer (P>0.01) and animal manure.
Microbial communities are major contributors of biodegradation of waste. Microbial load and diversity changes with the availability of organic matter during composting process. It is also influenced by the physicochemical conditions prevail throughout composting. In this study fungal community, their enzymatic capability, and the effect of abiotic parameters on their community during fruit waste composting were investigated using culture-dependent and culture independent microbiological techniques.
Starting with physicochemical parameters, temperature being a critical parameter during composting affects the waste degradation and its biological activity [24]. All phases of windrow composting in relation to temperature were observed within pile. The high temperature lasted for couple of days and then with progressive waste degradation it declined to mesophilic stage and then it reached to maturation stage where pile temperature remained unchanged with time to ambient temperature. These variation in temperature was the result of microbial metabolism which in turns favors shift in microbial community and was in accord with the previous studies [13,24,25].
Moisture content was gradually decreased over time despite of repetitive water addition. Since, water is required for microbes as a medium of nutrient transportation therefore, water loss indicates intense microbial activity resulting in heat generation that led to rise in temperature which eventually facilitates the decline in moisture content [2,26]. Several researchers [27–29] have also reported the same trend and claimed that gradual decrease in moisture content is indicative of stable and mature compost. The results showed that mature compost was stabilized at pH 6.0. In accordance with another study [26], volatilization of ammoniacal nitrogen and release of H+ ions during microbial nitrification process contribute to low pH. It has also been reported that pH is not an absolute indicator of compost maturity; it primarily depends upon the type of feedstock and method used for composting [30].
Electrical conductivity (EC) has great impact towards compost application to agricultural land; it could act as limiting factor during germination of seed and growth of plant [31]. The data in our study expressed several gradual fluctuations over. However, the observed values were never below the initial reading of conductivity. It has been reported that an upward trend of conductivity during composting process was observed due to release of ions like Ca2+, Mg2+, Na+, K+ and PO4-2 [29,32]. Likewise, the sudden decline in EC corroborated with another study where it was suggested that leaching of mineralized ions after water addition or formation of soluble salt complexes due to humification results in less number of free ions thereby a decline in EC is observed [25]. This idea has also been confirmed in a study of [33] where compost leachate has high conductivity. The finished compost has EC value < 3 mScm-1 indicating safe application of compost to soil [13].
C:N ratio not only constitute a significant parameter that indicates extent of complete biotransformation but also availability of nitrogen for plants [34]. Over the composting period, C:N ratio declined consistently and significantly (P < 0.05) and ended at 19:1 The drop in C:N ratio is relevant with the increase in N, which is because of mineralization of nitrogenous compounds, and decrease in C, due to production of CO2 during organic matter decomposition [31]. These findings are in consent with the findings of Carmo et al. [35] where a correlation in increase in inorganic matter loss with the exhaustion in C:N ratio was observed. A possible reason is the accelerated activity of microbial decomposition. In the present investigation, C:N ratio of mature compost was found to be in the optimum range i.e. <20:1 that can enable the availability and accessibility of nutrients for plants upon application [4].
Nutrient concentration of the final compost varies with the feedstock and method applied for decomposition [34]. In this study, major macronutrients; Phosphorus (P), Calcium (Ca) and Magnesium (Mg) were estimated. Individually, Ca and Mg showed escalation, as process progressed till the end and these observations are also in agreement with the reported results [13,36]. On the contrary, P content escalated (P < 0.05) on 7th day, remained static (P > 0.05) till day 76th and then took significant decline (P < 0.05) on 146th day. This dilution of nutrient during 76th day to 146th day of composting could be a result of leaching from the degraded waste [37]. The other possible explanation behind this sudden increase and decrease can be related to pH. As reported previously [31], Phosphorous content is very sensitive to pH; slight deviation from its optimum pH range i.e. 6-7 would result in hindering of its availability in soil. In agreement to this, Chandna et al. [36] has also correlated the solubility and stability of these minerals with pH of the mature compost.
To study the fungal community dynamics during fruit waste composting five different incubation temperatures with different incubation times were applied. This difference of incubation time between mesophilic and thermophilic fungal isolation was due to exposure of growth media to high temperature causing its dehydration. It was also observed that higher temperature favored quick growth of colonies in case of fungi. The mesophilic fungal load had increased not only with the passage of time but also with a rise in temperature whereas thermophilic fungal community underwent a decline even during thermophilic phase. Furthermore, as composting progressed towards the cooling stage, decrease in values of abiotic factors (pH, temperature, and moisture content) had resulted in high count of mesophilic fungi. In contrast to our study, Bhatia et al. [38] reported the decrease of fungal load with time during vegetable waste composting.
The shifts in microbial community structure were influenced by the nutritional factors in feedstock, local environmental factors and composting method used. The uniqueness in microbial population was due to the combinations of above-mentioned factors applied during composting process.
For fungi, a total of 50 isolates were preserved based on their incubation temperature and cultural characteristics. But out of this, only 35 (70 %) were categorized and their succession were explored. Remaining 30% of the isolates were excluded because of two reasons a) repetitive preservation of same morphotype and b) inability of their growth on sub-culturing into new vials. This sub-culturing inability into new vials was justified by López-González et al. [9] explaining the hindrance in isolation was occurred due to difference in nutrients, pH and specifically the competitive nature of microbes found in first culture media only. Reviewing the study done by Anastasia et al. [39], the results of fungal community are in agreement that recovered fungal isolates in our study are associated with the degradation of biowaste.
In matured compost sample, ITS amplicon based illumina Miseq platform revealed fungal diversity associated with three phyla including Zygomycota, Basidiomycota and Ascomycota and among phyla, Zygomycota was found as the most dominant one. Among Zygomycota, Mortierella sp. and Rhizomucor sp. were identified that are usually associated with decaying of the plant organic matters and appear in both mesophilic to thermophilic condition especially during composting of lignocellulosic materials [40]. Both species cause lignocellulose degradation and are mainly associated with hemicellulose and cellulose degradation [41]. However, most of Mortierella sp. and Rhizomucor sp. are usually termed as saprobe, ectomycorrhizal or plant pathogens that invade and take advantage to utilize soluble organic matters. Basidiomycota revealed as second dominant phylum, with Coprinopsis sp., Coprinellus canistri & Coprinus cordisporus. Species belong to class Agaricomycete are considered as efficient ligninolytic fungi with potential extracellular enzyme system. Most of these fungi can easily be seen on decaying wood with large fruiting bodies. They cause degradation of lignin first, leaving behind cellulosic material that usually utilize later [42]. Based on abundance, it can be understood that there must have been a gradual shift in wood-rotting fungi replaced with opportunist fungi i.e., zygomycetes. Results indicated less abundance 0.69 % of Ascomycota detected with Scutellinia sp., Schizothecium carpinicola, Penicillium sp., Preussia terricola, Arthrobotrys thaumasia, Zopfiellasp., Montagnulaceae sp., Trichoderma longibrachiatum, Aspergillus sp., Dactylellasp., Monascus purpureus, Myrmecridium schulzeri, Emericella sp. and Fusarium sp. Ascomycota are generally involved in soft decay of wood and show comparatively less potential of wood degradation than basidiomycetes. They are mainly attributed to hemicellulose and cellulose degradation [43]. The lowest abundance of ascomycetes can be linked with appearance of zygomycetes as opportunists on mature compost that attacked cellulosic material with highest abundance and deprived ascomycetes from its utilization.
In this study, fungal strains that were isolated during composting were screened for their enzymatic capabilities of degrading complex substances into simpler ones. Since, the compost pile has been made from fruit waste, leaves, and saw dust, it became a good source of pectin and cellulose, therefore, fungal strains were investigated for pectinolytic and cellulolytic activities.
Ten fungal isolates were found to be cellulase positive at 30 °C while only four fungal isolates were screened to be cellulase positive at 50 ºC, whereas three fungal isolates were found to be pectinase producers at mesophilic and thermophilic temperatures. These findings are in conformity with the findings of other studies that reported the capability of genera Aspergillus and Penicillium for the cellulase and pectinase production [19,44–47]. Overall, all lab designated isolate numbers (ADIF1, ADIB8 and ADIK2) of selected A. fumigatus were able to exhibit both cellulolytic and pectinolytic activity at thermophilic temperatures. Phutela et al. [48] has also reported the efficacy of A. fumigatus isolated from decomposing orange peel as a best producer of pectinase.
Culture-based analysis is not enough to give a true representation of microbial community in a rich environment like compost. It has also been reported that synthetic media has limited the species diversity and could only cultivate estimated < 1.5% of microbial diversity which shows biasness of culture-based results [18]. However, its significance should not be denied, and it must be coupled with the culture-independent techniques. Therefore, we opted a DGGE technique that uses DNA extracted from microbial community of compost sample enable us to have a better representation of diversified microbial community. Our study has revealed primary combination of sawdust and fruit waste provided a balanced nutrient and good moisture content to carryout composting process efficiently in a period of 146 days. The assessment of physicochemical parameters assured the maturity and quality of compost and its transition had influenced the microbial shifts during composting process. The periodic increase of fungal community with time indicates that mycoflora are secondary degraders that participate in decomposition of recalcitrant substrates. DGGE revealed that the fungal community is far more diverse than presented by culturing. Substrates availability, temperature and moisture content seemed to play a key role in designing the composition of fungal community, present at the respective stages of the composting process [49]. The decrease in the number of bands from week 0 to week 4 suggesting that the fungal specie cannot survive higher temperatures [7]. It has been reported in several studies that when the temperature of the compost pile, fell to around 40 °C, abundant fungal growth was apparent [3,50]. When the temperature drops, the fungal population starts recolonizing the compost [2,3]. Earlier investigation determined the relationship of physicochemical conditions and microbial population present in compost of municipal solid waste [51]. They reported that at ca.60 °C, the count of yeasts and filamentous fungi decreased from 106 to 103cells g-1, thus while unable to grow and causing loss in viability, significant numbers survived and were able to recolonize the compost once the temperature dropped. Cutting and sequencing of DGGE gel bands could enhance the resolution of DGGE, although the attempt failed in our case (data not shown).
Several studies using pyrosequencing, suggest that at thermophilic stage of composting process, selection of thermophilic and thermotolerant fungal population with decrease in the fungal diversity at this stage [2,7,52]. In the study, Zafar et al., [18] incubated compost at 45 °C and 50 °C for 12 weeks and reported that, Emericella rugulosa, and Scytallidium thermophilum accounted for 81 % and 88.9 % of all sequences at 50 °C and 45 °C respectively.
Genus Aspergillus was the most recovered fungi via culturing but according to the DGGE results it was sparsely present in compost. In a previous study using 454 pyrosequencing, A.fumigatus and T.lanuginosus accounted for <0.35% of the sequences, despite appearing to be the only organisms through conventional cultivation [7]. Culture based techniques depends upon the conditions of cultivation that favors the proliferation of fungi with less biomass from small spores number which is too small to be detected by PCR based methods [49]. DGGE is an important technique but like other molecular techniques, it has limitations too. Several reports are published reporting the limitations of DGGE e.g., co-migration of bands could underestimate while heterogeneity of genes could overestimate the diversity. Use of universal primers for amplification of gene could results in prevalence and ignorance of predominant and minor constituents of community, respectively [53–55].
As far as, phytotoxicity of mature compost is concerned, the study of Choy et al. [56] also suggested to use fruit peels for composting as it gives better maturity to compost and result in significant root growth of seeds.
Overall, our study has concluded that fruit waste is highly suitable for composting that produces enriched compost and indicates a future potential to be utilized as good organic fertilizer.
CFU, Colony forming unit; DGGE, Denaturing gradient gel electrophoresis; EC, Electrical conductivity; ITS, Internal transcribed spacer; IU, International unit; NGS, Next generation sequencing: PCR, Polymerase chain reaction; SDA, Sabourauds dextrose agar.
Funding: This study was financially supported by Higher Education Commission, Pakistan through Start-up Research Grant Program No:21-115/SRGP/R&D/ HEC/ 2014.
Conflict of Interest: The authors declare that there is no conflict of interests.
Availability of Data and Material: The authors declare that [the/all other] data supporting the findings of this study are available within the article [and its supplementary information files]
Code Availability: Not Applicable
Authors’ Contribution: Syeda Abeer Danish designed and planned the study, carried out all the experiments, performed analysis and interpretation of results and draft the manuscript. Urooj Zafar conceived the original idea and supervised the project. Tooba Haq performed the chemical analysis of compost samples. Mohammad Qureshi carried out catabolic profiling of fungal isolates. Iram Liaqat and Saima Rubab performed Next generation sequencing. All authors discussed the results and contributed to the final manuscript.
Ethics Approval: Not Applicable
Consent to Participate: Not applicable
Consent for Publication: All authors have agreed to publish article in your journal
ACKNOWLEDGEMENTS:
This study was financially supported by Higher Education Commission, Pakistan through Start-up Research Grant Program No:21-115/SRGP/R&D/ HEC/ 2014. The authors also thank Dr Muhammad Sohail, department of Microbiology, University of Karachi for his critical review on the manuscript.
- Kaza S, Yao LC, Bhada-Tata P, Van Woerden F. What a Waste 2.0: A Global Snapshot of Solid Waste Management to 2050. Washington, DC: World Bank; 2018.
- Ryckeboer J, Mergaert J, Vaes K, Klammer S, De Clercq D, Coosemans J, et al. A survey of bacteria and fungi occurring during composting and self-heating processes. Ann Microbiol 2003;53:349–410.
- Ryckeboer J, Mergaert J, Coosemans J, Deprins K, Swings J. Microbiological aspects of biowaste during composting in a monitored compost bin. J Appl Microbiol 2003;94:127–37.
- Day M, Shaw K. Biological, Chemical, and Physical Processes of Composting. In: Peter J. Stoffella BAK, editor. Compost Util. Hortic. Crop. Syst. First, Boca Raton: CRC Press; 2001, p. 17–50.
- Liu Y, Deng Y, Chen P, Duan M, Lin X, Zhang Y. Biodegradation analysis of polyvinyl alcohol during the compost burial course. J Basic Microbiol 2019;59:368–74.
- Ishii K, Takii S. Comparison of microbial communities in four different composting processes as evaluated by denaturing gradient gel electrophoresis analysis. JApplMicrobiol 2003;95:109–19.
- Langarica-Fuentes A, Zafar U, Heyworth A, Brown T, Fox G, Robson GD. Fungal succession in an in-vessel composting system characterized using 454 pyrosequencing. FEMS Microbiol Ecol 2014;88:296–308. https://doi.org/10.1111/1574-6941.12293.
- Xi B, He X, Dang Q, Yang T, Li M, Wang X, et al. Effect of multi-stage inoculation on the bacterial and fungal community structure during organic municipal solid wastes composting. Bioresour Technol 2015;196:399–405. https://doi.org/10.1016/j.biortech.2015.07.069.
- Lopez-Gonzalez JA, Suarez-Estrella F, Vargas-Garcia MC, Lopez MJ, Jurado MM, Moreno J. Dynamics of bacterial microbiota during lignocellulosic waste composting: Studies upon its structure, functionality and biodiversity. Bioresour Technol 2015;175:406–16.
- Dang Q, Wang Y, Xiong S, Yu H, Zhao X, Tan W, et al. Untangling the response of fungal community structure, composition and function in soil aggregate fractions to food waste compost addition. Sci Total Environ 2021;769:145248. https://doi.org/10.1016/j.scitotenv.2021.145248.
- Federation W. Standard Methods for the Examination of Water and Wastewater. Wahington D.C: American Public Health Association, American Water Works Association, Water Environment Federation; 1999.
- Langarica-Fuentes A, Fox G, Robson GD. Metabarcoding analysis of home composts reveals distinctive fungal communities with a high number of unassigned sequences. Microbiology 2015;161:1921–32. https://doi.org/10.1099/mic.0.000153.
- Haq T, Begum R, Ali TA, Iqbal S, Khan FA. Recycling of Biomass Waste from Herbal Pharmaceutical Industry by Windrow Composting. Int J Sci Res Publ 2016;6:257–62.
- Barratt SR, Ennos AR, Greenhalgh M, Robson GD, Handley PS. Fungi are the predominant micro-organisms responsible for degradation of soil-buried polyester polyurethane over a range of soil water holding capacities. J Appl Microbiol 2003;95:78–85.
- Feng J, Hwang R, Chang KF, Hwang SF, Strelkov SE, Gossen BD, et al. An inexpensive method for extraction of genomic DNA from fungal mycelia. Can J Plant Pathol 2010;32:396–401. https://doi.org/10.1080/07060661.2010.508620.
- White T. J., Bruns T. D., Lee S. B., . J. W. T. Amplification and direct sequencing of fungal ribosomal RNA Genes for Phylogenetics. PCR Protoc. A Guid. to Methods Appl., Academic Press, Inc; 1990, p. 315–22.
- Cosgrove L, McGeechan PL, Robson GD, Handley PS. Fungal communities associated with degradation of polyester polyurethane in soil. Appl Environ Microbiol 2007;73:5817–24.
- Zafar U. Biodegradation of Polyurethane under Composting Conditions : A thesis submitted to The University of Manchester for Faculty of Life Sciences Urooj Zafar. 2013.
- Sohail M, Siddiqi R, Ahmad A, Khan SA. Cellulase production from Aspergillus niger MS82: effect of temperature and pH. N Biotechnol 2009;25:437–41. https://doi.org/10.1016/j.nbt.2009.02.002.
- Khokhar I, Haider MS, Mushtaq S, Mushtaq I. Isolation and Screening of Highly Cellulolytic Filamentous Fungi. J Appl Sci Environ Manag 2012;16:223–6.
- Ahmed A, Ejaz U, Sohail M. Pectinase production from immobilized and free cells of Geotrichum candidum AA15 in galacturonic acid and sugars containing medium. J King Saud Univ - Sci 2020;32:952–4. https://doi.org/10.1016/j.jksus.2019.07.003.
- Ansari M, Zafar U, Ejaz U, Sohail M, Pirzada A, Aman A. Comparison of composting of chemically pretreated and fermented sugarcane bagasse for zero-waste biorefinery. J Mater Cycles Waste Manag 2021. https://doi.org/10.1007/s10163-021-01176-w.
- Ahmad R, Jilani G, Arshad M, Zahir ZA, Khalid A. Bio-conversion of organic wastes for their recycling in agriculture: an overview of perspectives and prospects. Ann Microbiol 2007;57:471–9.
- Wei H, Wang L, Hassan M, Xie B. Succession of the functional microbial communities and the metabolic functions in maize straw composting process. Bioresour Technol 2018;256:333–41. https://doi.org/10.1016/j.biortech.2018.02.050.
- Mengistu T, Gebrekidan H, Kibret K, Woldetsadik K, Shimelis B, Yadav H. Comparative effectiveness of different composting methods on the stabilization, maturation and sanitization of municipal organic solid wastes and dried faecal sludge mixtures. Environ Syst Res 2018;6:5.
- Dehghani R, Asadi MA, Charkhloo E, Mostafaie G, Saffari M, Mousavi GA, et al. Identification of Fungal Communities in Producing Compost by Windrow Method. J Environ Prot (Irvine, Calif) 2012;03:61–7.
- Ameen A, Ahmad J, Raza S. Effect of pH and moisture content on composting of Municipal solid waste 2016;6:35–7.
- Nasreen Z, Qazi JI. Lab scale composting of fruits and vegetable waste at elevated temperature and forced aeration. Pak J Zool 2012;44:1285–90.
- Fernandez-Delgado Juarez M, Pr??hauser B, Walter A, Insam H, Franke-Whittle IH. Co-composting of biowaste and wood ash, influence on a microbially driven-process. Waste Manag 2015;46:155–64. https://doi.org/10.1016/j.wasman.2015.09.015.
- Azim K, Soudi B, Boukhari S, Perissol C, Roussos S, Thami Alami I. Composting parameters and compost quality: a literature review. Org Agric 2017;8:141–58. https://doi.org/10.1007/s13165-017-0180-z.
- Ashman M, Puri G. Essential Soil Science : a Clear and Concise Introduction to Soil Science. John Wiley & Sons; 2013.
- Shemekite F, Gomez-Brandon M, Franke-Whittle IH, Praehauser B, Insam H, Assefa F. Coffee husk composting: An investigation of the process using molecular and non-molecular tools. Waste Manag 2014;34:642–52.
- Mohammadipour Z, Enayatizamir N, Ghezelbash G, Moezzi A. Bacterial Diversity and Chemical Properties of Wheat Straw-Based Compost Leachate and Screening of Cellulase Producing Bacteria. Waste and Biomass Valorization 2020.
- Kokkora MI, Hann MJ, Tyrrel SF. Organic Waste Compost Parameters in Relation To Soil Properties. Proc Int Agric Eng 2008 Conf Ind Exhib 23 25 June 2008 2008.
- Carmo DL do, Lima LB de, Silva CA. Soil fertility and electrical conductivity affected by organic waste rates and nutrient inputs. Rev Bras Ciência Do Solo 2016;40:1–17.
- Chandna P, Nain L, Singh S, Kuhad RC. Assessment of Bacterial Diversity during Composting of Agricultural Byproducts. BMC Microbiol 2013;13:99.
- Jakubus M. Estimation of Phosphorus Bioavailability from Composted Organic Wastes. Chem Speciat Bioavailab 2016;28:189–98.
- Bhatia A, Ali M, Sahoo J, Madan S, Pathania R, Ahmed N, et al. Microbial diversity during Rotary Drum and Windrow Pile composting. J Basic Microbiol 2012;52:5–15. https://doi.org/10.1002/jobm.201100077.
- Anastasi A, Varese GC, Marchisio VF. Isolation and identification of fungal communities in compost and vermicompost. Mycologia 2005;97:33–44.
- Busk PK, Lange L. Cellulolytic Potential of Thermophilic Species from Four Fungal Orders. AMB Express 2013;3:1–10.
- Andlar M, Rezić T, Marđetko N, Kracher D, Ludwig R, Šantek B. Lignocellulose degradation: An overview of fungi and fungal enzymes involved in lignocellulose degradation. Eng Life Sci 2018;18:768–78. https://doi.org/10.1002/elsc.201800039.
- Badalyan SM, Szafranski K, Hoegger PJ, Navarro-González M, Majcherczyk A, Kües U. New Armenian wood-associated coprinoid mushrooms: Coprinopsis strossmayeri and Coprinellus aff. radians. Diversity 2011;3:136–54. https://doi.org/10.3390/d3010136.
- Mäkelä MR, Marinović M, Nousiainen P, Liwanag AJM, Benoit I, Sipilä J, et al. Aromatic metabolism of filamentous fungi in relation to the presence of aromatic compounds in plant biomass. Adv Appl Microbiol 2015;91:63–137. https://doi.org/10.1016/bs.aambs.2014.12.001.
- Priya V, Sashi V. Pectinase Enzyme Producing Microorganisms. Int J Sci Res Publ 2014;4.
- Sohail M, Ahmad A, Khan SA. Production of cellulase from Aspergillus terreus MS105 on crude and commercially purified substrates. 3 Biotech 2016;6:1–8. https://doi.org/10.1007/s13205-016-0420-z.
- Villena GK, Gutiérrez-Correa M. Production of cellulase by Aspergillus niger biofilms developed on polyester cloth. Lett Appl Microbiol 2006;43:262–8.
- Martínez-Trujillo A, Arreguín-Rangel L, García-Rivero M, Aguilar-Osorio G. Use of fruit residues for pectinase production by Aspergillus flavipes FP-500 and Aspergillus terreus FP-370. Lett Appl Microbiol 2011;53:202–9. https://doi.org/10.1111/j.1472-765X.2011.03096.x.
- Phutela U, Dhuna V, Sandhu S, Chadha BS. Pectinase and polygalacturonase production by a thermophilic Aspergillus fumigatus isolated from decomposting orange peels. Brazilian J Microbiol 2005:63–9.
- Cahyani VR, Matsuya K, Asakawa S, Kimura M. Succession and phylogenetic profile of eukaryotic communities in the composting process of rice straw estimated by PCR-DGGE analysis. Biol Fertil Soils 2004;40:334–44. https://doi.org/10.1007/s00374-004-0783-x.
- Finstein MS, Morris ML. Microbiology of Municipal Solid Waste Composting. Adv Appl Microbiol 1975;19:113–51.
- Hassen A, Belguith K, Jedidi N, Cherif A, Cherif M, Boudabous A. Microbial characterization during composting of municipal solid waste. Bioresour Technol 2001;80:217–25.
- Zafar U, Houlden A, Robson GD. Fungal Communities Associated with the Biodegradation of Polyester Polyurethane Buried under Compost at Different Temperatures. Appl Environ Microbiol 2013;79:7313–24. https://doi.org/10.1128/AEM.02536-13.
- Kang YJ, Cheng J, Mei LJ, Hu J, Piao Z YS. Multiple copies of 16s rRNA gene affect the restriction patterns and DGGE profile as revealed by analysis of genome database. Mikrobiologiia 2010;79:664–71.
- Jackson CR, Roden EE, Perry F. Denaturing Gradient Gel Electrophoresis Can Fail to Separate 16S rDNA Fragments with Multiple Base Differences 2000;1:49–51.
- Adil E. Corrective measures of denaturing gradient gel electrophoresis limitations. J Environ Sci Technol 2015;8:1–12. https://doi.org/10.3923/jest.2015.1.12.
- Choy SY, Wang K, Qi W, Wang B, Chen C-L, Wang J-Y. Co-composting of horticultural waste with fruit peels, food waste, and soybean residues. Environ Technol 2015;36:1448–56. https://doi.org/10.1080/09593330.2014.993728.
TABLE 1 Changes in Chemical parameters during Composting
|
Day of sample collection |
Total carbon (%) |
Total nitrogen (%) |
Carbon to Nitrogen ratio |
Macronutrients |
||
|
P (%) |
Ca (gkg-1) |
Mg (gkg-1) |
||||
|
0 |
21.00 |
0.68 |
30.61 |
1.14 |
19.67 |
9.67 |
|
7 |
19.67 |
0.87 |
22.52 |
1.37 |
50.33 |
15.33 |
|
76 |
20.00 |
0.88 |
22.72 |
1.36 |
52.67 |
18.33 |
|
146 |
17.67 |
0.90 |
19.49 |
1.13 |
58.00 |
17.67 |
TABLE 2 Catabolic profiling of selected fungal isolates. Qualitative and quantitative screening results of cellulase are presented. In case of Pectinase only qualitative screening was performed. + indicates enzyme production, - indicates no enzyme production, n/d indicates not determined
|
Lab Designated Number of Representatives |
Cellulase (IU ml-1) |
Pectinase |
||||
|
|
30 ºC |
50 ºC |
30 ºC |
50 ºC |
||
|
ADIF4 |
+ |
2.86 |
- |
- |
+ |
- |
|
ADIF9 |
+ |
3.01 |
- |
- |
+ |
- |
|
ADIB9 |
- |
- |
+ |
3.60 |
- |
- |
|
ADIF1 |
+ |
n/d |
+ |
4.40 |
- |
+ |
|
ADIB8 |
+ |
n/d |
+ |
3.56 |
- |
+ |
|
ADIK2 |
+ |
n/d |
+ |
3.39 |
- |
+ |
|
ADI8 |
+ |
1.12 |
- |
- |
- |
- |
|
ADIB3 |
+ |
1.62 |
- |
- |
+ |
- |
|
ADI7 |
+ |
n/d |
- |
- |
- |
- |
|
ADIB4 |
+ |
2.77 |
- |
- |
- |
- |
TABLE 3 Comparison of fruit waste composts with composted cattle manure and chemical fertilizer on the basis of germination Index of Vigna radiata.
|
Test samples used |
Aqueous extract Sample to water ratio (w/v) |
Germination Index (GI) % |
|
Fruit waste compost |
1:10 |
83.3 |
|
composted cattle manure |
1:10 |
70.4 |
|
chemical fertilizer |
0.25% of 1:10 of extract |
27.3 |
{kind=link}