Cloning of LhMYB12-Lat promoter
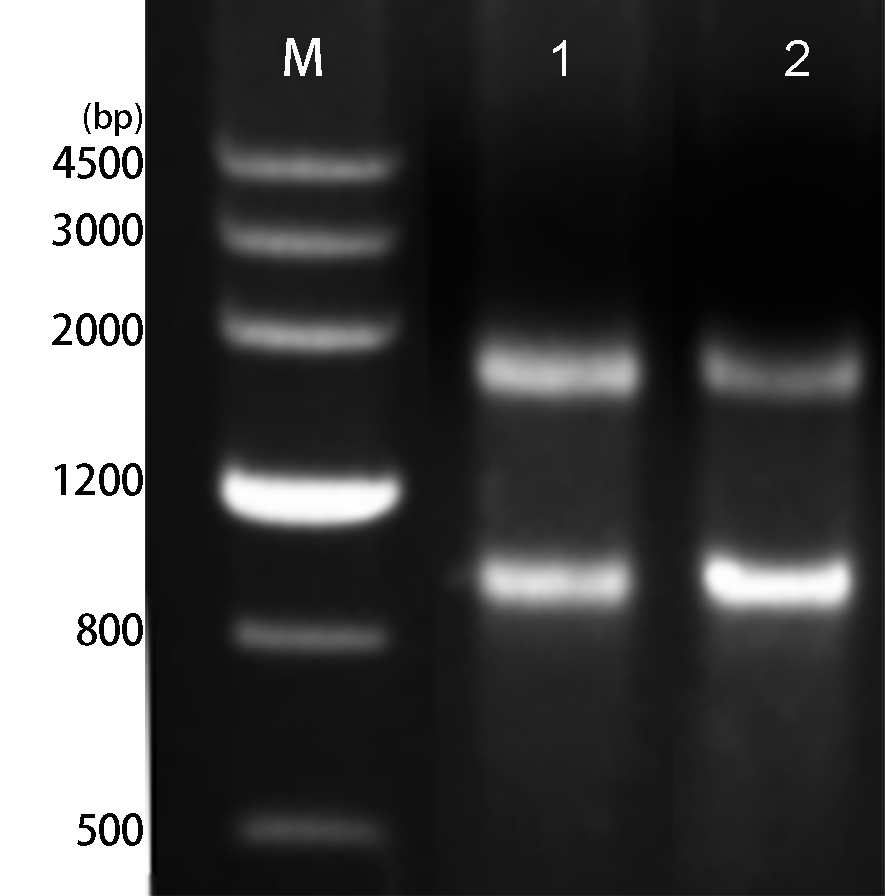
The lily LhMYB12-Lat promoter was isolated based on amplifying promoter sequences primers using the genome walking method and submitted as MW719044. The PCR products, a 3222 bp length, were verified by agarose gel electrophoresis (Fig. 1a) and shown in Fig.1b. The cis-elements and transcription factor binding sites showed ABA-, auxin-, MeJA-, gibberellin-, ethylene-, light-, and stress-responsive elements by online software PlantCARE and PlantPAN 2.0 in LhMYB12-Lat promoter sequence (Table 1).
GUS activity assay
To verify whether the promoter of LhMYB12-Lat could impact the expression of the LhMYB12-Lat gene, we constructed proLhMYB12-Lat::GUS recombination plasmids to infect the disc of lily tepals. The promoter activity can be directly reflected by GUS staining. As shown in Fig. 1c, GUS staining results showed that the LhMYB12-Lat promoter could activate gene expression of LhMYB12-Lat in lily tepals.
Table 1
Promoter cis-elements prediction.
|
Element
|
Sequence
|
Position
|
Function
|
|
A-box
|
CCGTCC
|
-912 to -907
|
cis-acting regulatory element
|
|
ABRE
|
ACGTG
|
-2726 to -2722
|
ABA-responsive element
|
|
ATCT-motif
|
AATCTAATCC
|
-3154 to -3144
|
part of a conserved DNA module involved in light-responsiveness
|
|
ARR
|
CAAATCT
|
-447 to -441
|
ARR motif
|
|
Box II
|
ACACGTAGA
|
-1882 to -1874
|
part of a light-responsive element
|
|
Box 4
|
ATTAAT
|
-216 to -211
|
light-responsive element
|
|
CAAT-box
|
CAAT
|
-98 to -95
|
common cis-acting element in promoter and enhancer regions
|
|
CAT-box
|
GCCACT
|
-935 to -930
|
cis-acting regulatory element related to meristem expression
|
|
CGTCA-motif
|
CGTCA
|
-1587 to -1583
|
cis-acting regulatory element involved in the MeJA-responsiveness
|
|
G-box
|
CACGTG
|
-1513 to -1508
|
cis-acting regulatory element involved in light-responsiveness
|
|
GATA-motif
|
GATAGGA
|
-198 to -192
|
part of a light-responsive element
|
|
GC-motif
|
CCCCCG
|
-1107 to -1102
|
enhancer-like element involved in anoxic specific inducibility
|
|
GCC-box
|
CCGCCGTC
|
-376 to -369
|
ethylene-responsive element
|
|
GCN4_motif
|
TGAGTCA
|
-821 to -815
|
cis-regulatory element involved in endosperm expression
|
|
GT1-motif
|
GGTTAA
|
-507 to -502
|
light-responsive element
|
|
HSE
|
AGAANNTTCT
|
-2869 to -2861
|
HSF element
|
|
LAMP-element
|
CTTTATCA
|
-2467 to -2460
|
part of a light-responsive element
|
|
LTR
|
CCGAAA
|
-792 to -787
|
cis-acting element involved in low-temperature responsiveness
|
|
MBS
|
CAACTG
|
-3091 to -3086
|
MYB binding site involved in drought-inducibility
|
|
Myb
|
CAACTG
|
-470 to -465
|
Myb motif
|
|
MYC
|
CATGTG
|
-2540 to -2539
|
MYC element
|
|
P-box
|
CCTTTTG
|
-3005 to -2999
|
gibberellin-responsive element
|
|
Sp1
|
GGGCGG
|
-1673 to -1668
|
light-responsive element
|
|
TATA-box
|
TATA
|
-30 to -27
|
core promoter element around -30 of transcription start
|
|
TCA-element
|
CCATCTTTTT
|
-1896 to -1887
|
cis-acting element involved in salicylic acid responsiveness
|
|
TCCC-motif
|
TCTCCCT
|
-1116 to -1110
|
part of a light-responsive element
|
|
TGA-element
|
AACGAC
|
-1818 to -1813
|
auxin-responsive element
|
|
TGACG-motif
|
TGACG
|
-1587 to -1583
|
cis-acting regulatory element involved in the MeJA-responsiveness
|
|
W box
|
TTGACC
|
-2561 to -2556
|
WRKY motif
|
Construction of a cDNA library
The Asiatic hybrid lily ‘Tiny Padhye’ upper tepals and bases of tepals from the S2 stage were collected for total RNA extraction. The quality of the total RNA samples is shown in Fig. S1, showing the bands corresponding to the intact 28S and 18S rRNA. The total RNAs had an A260/A280 ratio of 2.06-2.10 and a concentration of 480-556 ng/μL, which fulfilled the conditions for constructing a cDNA library.
Agarose gel electrophoresis showed that the cDNA library was successfully constructed, and the primary library volume was above 1.6Í106 CFU, with a recombination efficiency of more than 98%. The homogenization results verified that the redundancy rate was 1% by sequencing 96 single positive clones (Fig. 2).
Construction of bait-reporter strains and determining AbA concentration
As shown in Fig. 3a, the colony PCR results were consistent with the expected size of PCR products (1.35 kb plus the inserted fragment), identifying correct integration into yeast cells.
To exclude yeast transcription factors for target sequence recognition, we measured minimum concentrations of AbA for inhibiting self-activation. As shown in Fig .3b, basal expression of pAbAi-LhMYB12-Lat-P1(elements), pAbAi-LhMYB12-Lat-P2 and pAbAi-LhMYB12-Lat-P3 bait strains were 250 ng/mL, 500 ng/mL, and 500 ng/mL, respectively (Fig. 3c).
Screening of a yeast one-hybrid library
pAbAi-LhMYB12-Lat-P1(elements), pAbAi-LhMYB12-Lat-P2, and pAbAi-LhMYB12-Lat-P3 yeast cell suspensions obtained on SD/-Leu plates were diluted to 1/100, and 124, 77, and 20 clones, respectively. The number of screened clones were 1.9Í106, 1.2Í106, and 0.3Í106, respectively.
Extraction of the prey plasmids and confirming positive interactions
The positive colonies of pAbAi-LhMYB12-Lat-P1(elements), pAbAi-LhMYB12-Lat-P2 and pAbAi-LhMYB12-Lat-P3 were selected from the SD/-Leu plates, 149, 140, and 141 clones, respectively, and grow on SD/-Leu brown media to extract the prey plasmids. Then, the extracted plasmids were transferred with prey strains to the SD/-Leu medium containing the appropriate concentration of AbA, and the pGADT7 plasmid was used as a negative control. The positive plasmids of the growing yeast strains were identified that regulate the expression of the LhMYB12-Lat gene by interacting with cis-elements in the LhMYB12-Lat promoter. Finally, the confirmed sequences of lily were compared by Blastx (https://blast.ncbi.nlm.nih.gov/Blast.cgi), including Zinc finger, ERF, GRP, and MYB predicted proteins, which originate from Elaeis guineensis, Phoenix dactylifera, and Musa acuminata (Table 2).
Expression analysis of candidate TFs of LhMYB12-Lat
To verify the interaction between the expression levels of candidate TFs and LhMYB12-Lat, we analyzed the expression of candidate TFs genes in ‘Tiny Padhye’ tepals by qRT-PCR. We found that expression levels of most genes tested were significantly different in the upper and basal tepals (Fig. 4a). These results suggest that the genes (MYB4, ERF WIN1-like, ERF061, ERF071-like, ARR1, BTF3, IAA6-like, and ERF4) may be involved in the suppression of LhMYB12-Lat-mediated anthocyanin biosynthesis regulation in lily, and that HSF, ZF CCCH and GRP6 genes may determine the positive or negative effect on LhMYB12-Lat.
Table 2
Screening results of LhMYB12-Lat promotor transcriptional factors.
|
Accession number
|
Biological annotation
|
Species
|
|
AEU17861.1
|
heat shock transcription factor
|
Lilium longiflorum
|
|
XP_012476945.1
|
basic transcription factor 3
|
Gossypium raimondii
|
|
ASV46333.1
|
MYB4
|
Lilium regale
|
|
RWR72505.1
|
WD-40 repeat
|
Cinnamomum micranthum
|
|
XP_010913061.1
|
zinc finger CCCH domain-containing protein ZFN-like isoform X2
|
Elaeis guineensis
|
|
XP_008810485.1
|
auxin-responsive protein IAA6-like isoform X1
|
Phoenix dactylifera
|
|
XP_009412068.1
|
ethylene-responsive transcription factor 4
|
Musa acuminata
|
|
XP_009387936.1
|
ethylene-responsive transcription factor ERF071-like
|
Musa acuminata
|
|
XP_030477126.1
|
gibberellin-regulated protein 6
|
Syzygium oleosum
|
|
XP_010905671.1
|
two-component response regulator ARR1
|
Elaeis guineensis
|
|
XP_008804592.1
|
ethylene-responsive transcription factor WIN1-like
|
Phoenix dactylifera
|
|
XP_010913767.1
|
ethylene-responsive transcription factor ERF061
|
Elaeis guineensis
|
LUC reporter assay
we performed LUC reporter assays to determine how several potential TF genes regulate the activity levels of proLhMYB12-Lat. As shown in Fig. 4b-c, BTF3, IAA6-like, ERF4, ERF WIN1-like, ERF061 negative regulated proLhMYB12-Lat, but MYB4 and ARR1 barely affected the activity level of proLhMYB12-Lat. Thus, BTF3, IAA6-like, ERF4, ERF WIN1-like, ERF061 was found to mainly suppress proLhMYB12-Lat to negative regulate anthocyanin accumulation in lily.
{kind=link}