LATS1 predicts poor clinical outcomes and knockdown of LATS1 inhibits the proliferation of BLCA cells in vitro and in vivo.
To explore the possible biological functions of LATS1 in BLCA, first we evaluated the association between LATS1 expression of and patients’ prognosis in BLCA using TIMER database. Kaplan-Meier analysis showed that BLCA patients with high LATS1 expression had shorter overall survival (OS) (Figure 1A). According to multivariate Cox regression analyses using BLCA data from the TCGA bladder cancer data set, LATS1 expression might be a significant prognostic factor for reduced OS (HR = 1.89, P =0.001) and progression free survival (PFS) (HR = 1.75, P = 0.013) (Figure 1B and 1C), which suggests that high expression of LATS1 is a predictor of poor prognosis in BLCA patients.
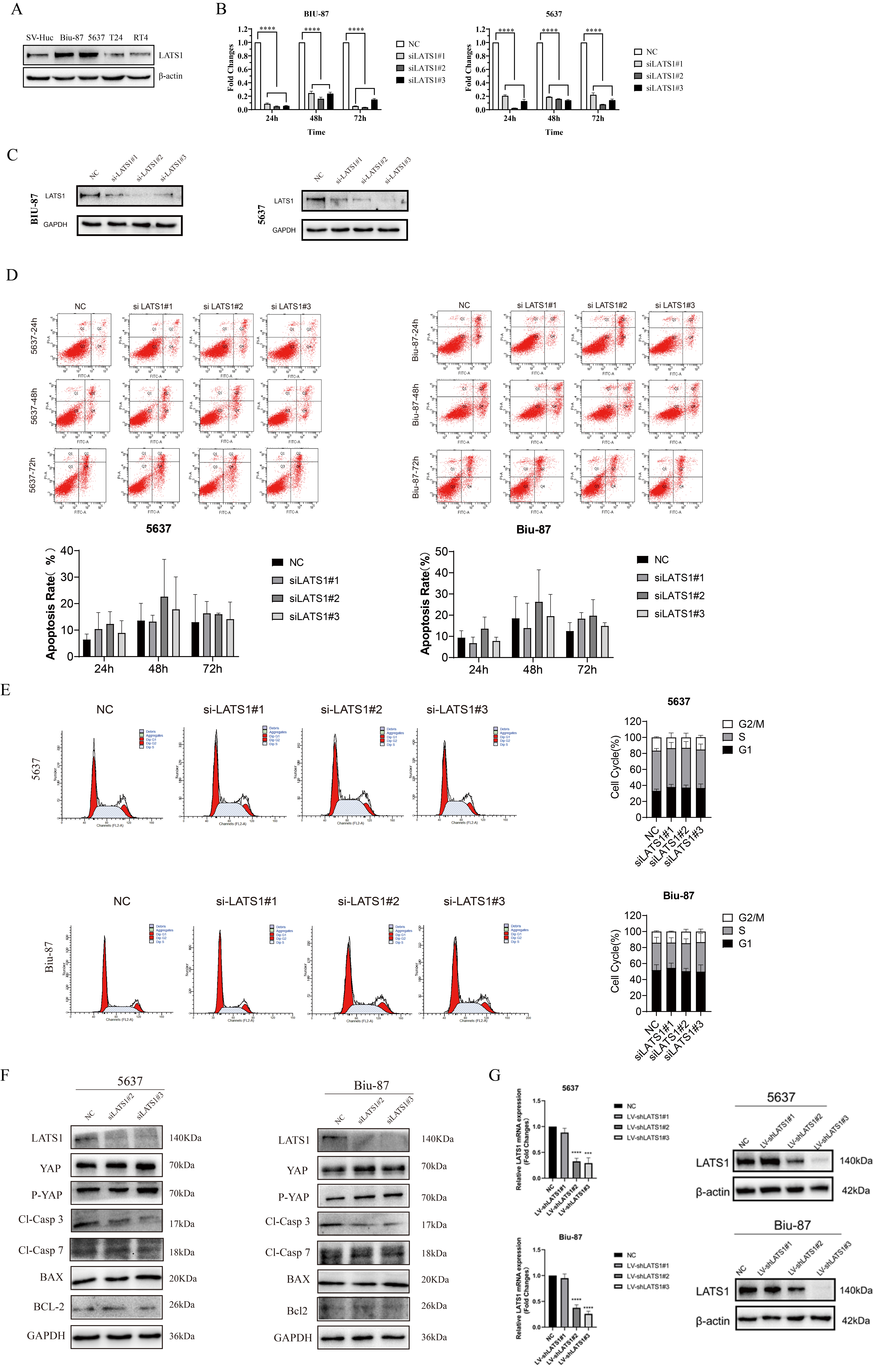
To further confirm the function of LATS1 in BLCA, we examined LATS1 expression by western blot analysis in BLCA cell lines (Figure S1A). And then, BIU-87 and 5637 cells with higher LATS1 expression were selected to assess the effects of LATS1 changes on the cell proliferation in vitro for this study. Then, small interfering RNA (siRNA) was used to down-regulate LATS1 in BIU-87 and 5637 cells (Figure S1B and S1C). siRNA-mediated knockdown of LATS1 (si-LATS1) resulted in a statistically significant reduction of cell viability and a decrease of colony formation capacity in different BLCA cell lines (Figure 1D and 1E). Interestingly, analysis of apoptosis by Flow Cytometry showed that the number of apoptotic cells was not significantly increased in si-LATS1 cells compared with non-targeting control (NC) cells (Figure S1D), and the western blot analysis showed that the protein expressions of cleaved caspase 3, 7, BAX, BCL-2 were not significantly changed in si-LATS1 cells (Figure S1E). To confirm whether si-LATS1 affected its classic target YAP1, we examined the expression of YAP1 and phospho-YAP1 with Western Blot. We found that there were no significant changes of both YAP1 and phospho-YAP1 in the si-LATS1 cells than that in NC cells (Figure S1E). Moreover, si-LATS1 did not cause significant cell cycle change (Figure S1F), too.
For the in-vivo study, lentiviral-delivered shRNAs were used to inhibit LATS1 expression (Figure S1G) in 5637 cells to obtain LATS1 knockdown cell line after screened by neomycin. And then, cell-line-derived tumor xenograft (CDTX) model was established by subcutaneously injecting 5x106 5637 cells infected with empty lentivirus vector or lentivirus LATS1 shRNAs. The BLCA cell tumor began to be measurable 14 days after cell injection in the control group (LV-vector group), but there was no significant subcutaneous BLCA cell tumor found in LV-shLATS1 groups (Figure 1G).
Together, these findings demonstrated that LATS1 could serve as a prognostic predictor of BLCA survival, and si-LATS1 could inhibit the proliferation of BLCA cells in vitro and in-vivo, but without induction of cell apoptosis and cell cycle arrest. And Hippo signaling was not involved in the proliferation suppression induced by si-LATS1 in BLCA cells.
Gene enrichment analysis of LATS1 knockdown and wild-type BLCA cells
To elucidate the potential genes and possible pathways affected by knockdown of LATS1, BIU-87 cells were transfected with siRNA or siNC. After 24 hours, the RNA was isolated from the siRNA andsiNC cells for use in Bulk RNA-sequencing (RNA-seq). Based on the R analysis, a total of 715 differentially expressed genes (DEGs), 501 upregulated genes and 214 downregulated genes, were identified in the cells of si-LATS1 groups compared with NC groups (Figure 2A). P<0.05 and |log2 FC|≥1were set as the threshold criteria.
To further explore the systematic characterization and biological functions of the identified DEGs, functional annotation and pathway analysis, including GO and KEGG, were performed using R packages.In this study, the twoGO categories [biological process (BP) and molecular function MF)] were detected, respectively, using R packages. The GO analysis showed that DEGs were significantly enriched in different biological processes (BP) and molecular functions (MF), such as “response to lipopolysaccharide”(ontology: BP) and “DNA-binding transcription activator activity” (ontology: MF) (Figure 2B). Subsequently, KEGG pathway analysis showed that the DEGs were enriched in several key pathways, including ‘TNF signaling pathway’,‘Rheumatoid arthritis’,and ‘NF-kappa B signaling pathway’ (Figure 2C).
Using the RNA-seq data, we also performed gene set enrichment analysis (GSEA) to extract cellular functions associated with LATS1 knockdown. GSEA revealed that the changes in gene expression caused by the knockdown of LATS1 correlated positively with high expression of inflammatory genes, especially HALLMARK_TNFA_SIGNALING_VIA_NFKB regulated genes (Figure 2D) (total 14 gene sets weremeaningful, P<0.05, FDR<0.05) from ‘h.all.v7.4.symbols gene set collection’, and REACTOME_INFLAMMASOMES (total 16 gene sets weremeaningful, P<0.05, FDR<0.05) from ‘c2.cp.reactome.v7.4symbols gene set collection’.
Through the bioinformatic analyses above, we found that the alterations in gene expression caused by si-LATS1 were most related with inflammatory responses and pathways. Generally, all the BLCA cells in this study were harvested at 24h unless otherwise specified.
LATS1 knockdown increasesthe expressions of pro-inflammatory cytokinesvia activating TNF-α/NF-κB signalingin BLCA cells
Based on the comprehensive bioinformatic analysis above, we first focused on the TNFA_SIGNALING_VIA_NFKBpathway (Figure 3A). To validate whether si-LATS1 induces the inflammatory responses in the BLCA cells, we used RT-qPCR to detect the mRNA expressions of some pro- and anti-inflammatory cytokine genes, most of which are the targets of NF-κB. We found that si-LATS1 could significantly increase the mRNA expressions of TNF-α, IL-1β, IL-18 (Figure 3B), IFN-γ,IL-2 and IL-6 (Figure S2A-2B) but decrease the expressions of IL-10, TGF-β and IL-4 (Figure S2C-2F). The Western Blot results demonstrated that the expression of phospho-NF-κB/RelA and secreted TNF-α(sTNF-α) in si-LATS1 cells was significantly higher than that in their respective control (NC) cells, but without changing the expression of totalNF-κB/RelA (Figure 3C).Immunofluorescence (IF) results confirmed that si-LATS1 could promote the translocation of NF-κB from cytoplasm to nucleus (Figure 3D).
To further determine if NF-κB activation was required for inflammation responses in si-LATS1 cells, the NF-κB inhibitor PS341 was used when the BLCA cells were transfected with LATS1 siRNAs. We found that the mRNA expressions of TNF-α, IL-1β, IL-6 and IL-18 were obviously downregulated in the si-LATS1 cells co-treated with 200nM PS341 compared with those in the si-LATS1 cells (Figure S2G; Figure 3E).
All the results above proved that si-LATS1 could induce the activation TNF-α/NF-κB signaling,increasing the expressions of pro-inflammatory cytokines in BLCA cells.
LATS1 knockdown induces cellular pyroptosis via TNF-α/NF-κB signaling
To further confirm thenon-apoptotic way of the cell death, DNA ladder assay was performed using agarose gel electrophoresistoassess DNA fragmentation in the BLCA cells transfected with LATS1 siRNA. We found that DNA cleavageswere random and non-specific in the si-LATS1 cells (Figure 4A), which suggested that the cell death was predominantly non-apoptotic. Additionally, lactate dehydrogenase (LDH)-release into the culture medium was quantified to estimate the plasma membrane damagein si-LATS1 cells. And the results showed that the levels of LDH in culture medium were significantly elevated in the si-LATS1 cells compared with those in the NC cells (Figure 4B).
Based on the previous results and the bioinformatic analysis, we focused on the INFLAMMASOMEpathway (Figure 4C and Figure 2E). The genes retrieved from the gene set (REACTOME_INFLAMMASOMES)were calculated by the cytoHubba in Cytoscape software according to the degree of connectivityto identify the possible hub genes that si-LATS1 affected, and the top 10 genes in protein-protein interaction (PPI) network were considered as hub genes (Figure 4C and 4D).
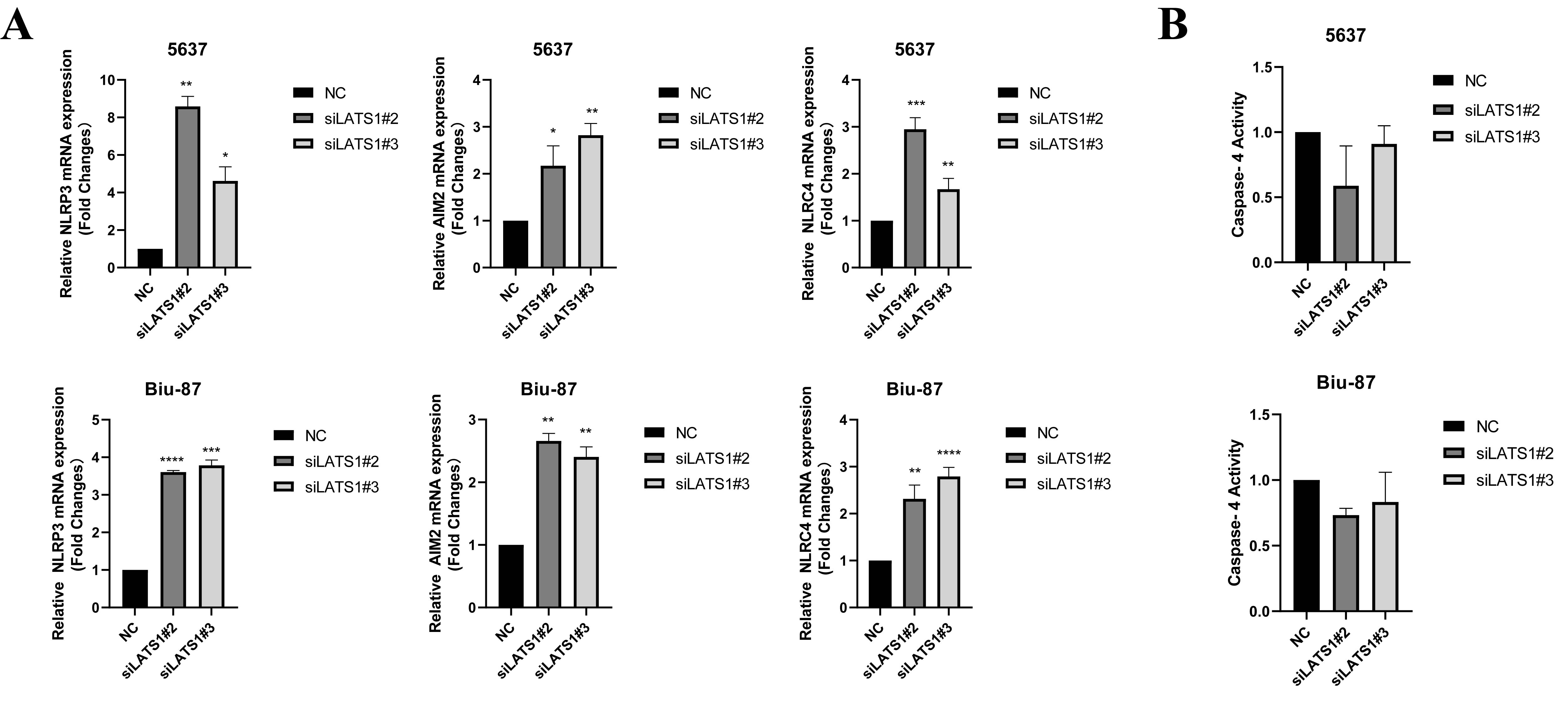
Up to date, several inflammasomes have been identified, including AIM2, NLRP3 and NLRC4, which have been reported to protect against tumor growth(38). We found that the expressions of AIM2, NLRP3 and NLRC4 were obviously elevated in the si-LATS1 cells at mRNA (Figure S3A) and protein (Figure 4F) levels.In the meantime, the activity of caspase 1 [but not caspase 4 (Figure S3B)] was significantly increased in the si-LATS1 cells than that in the NC cells (Figure 4E and 4F); and the expressions of IL-1β and IL-18 were significantly increased; as the executioner of cell pyroptosis, GSDMD was cleaved and activated (Figure 4F). The addition of pan-caspase inhibitor Z-VAD-FMK markedly rescued the si-LATS1-induced cell viability reduction in the BLCA cells (Figure 4G). Furthermore, the addition of the NF-κB inhibitor PS341 also reduced the upregulated expressions of AIM2, NLRP3 and NLRC4 in the si-LATS1 cells (Figure 4H).
Taken together, si-LATS1 contributed to the formation of AIM2, NLRP3 and NLRC4 inflammasomes via the activation of TNF-α/NF-κB signaling, inducingthe cellular pyroptosis in the BLCA cells.
LATS1 overexpression inhibitsTNF-α/NF-κB and the inflammasome pathways
To investigate roles of overexpressed LATS1 (OE-LATS1) in the activation of TNF-α/NF-κB signaling and the formation ofinflammasomes, we transfected LATS1-expressing plasmid into BIU-87 and 5637 cells. And the expression of LATS1 was obviously enhanced in BLCA cells after transfection with human the LATS1 plasmid at mRNA and protein levels (Figure 5A). And we found that the cellular viabilities of BIU-87 and 5637 cells were not evidently affected in the OE-LATS1 cells compared with NC cells (Figure 5B). But the expressions of sTNF-α and phospho-NF-κB/RelA were markedly downregulated (Figure 5C). And the protein expressions of AIM2, NLRP3 and NLRC4, cleaved caspase 1, cleaved GSDMD were all clearly decreased in the OE-LATS1 cells (Figure 5D). In addition, the overexpression of the LATS1 has been shown to rescue the proliferation in the si-LATS1 cells (Figure 5E).
The results showed that LATS1 overexpression inhibited TNF-α/NF-κB signaling to block the formation ofAIM2, NLRP3 and NLRC4 inflammasomes.
LATS1 knockdown induced pryoptosis of BLCA cells promotes M1 polarization of THP1 derived macrophages
TAMs are important components of theTME, which can beused as the therapeutic targets of BLCA treatment.First, we used xCell R package to analyze thetumor‑infiltrating immune cell in BLCAtissues using the BLCA data obtained from TCGA database.The results indicated that infiltration levels of CD8+ T cells, B cells, myocytes, NK cells,pDC,plasmacells, and smooth muscle cells were significantly associated withthe overall survival of BLCA patients, including the M2 macrophages (Figure 6A).Tovalidate the effects of si-LATS1 induced pryoptosis on the polarization of macrophages, THP-1 monocytes were polarized into macrophages using PMAandthen PMA-treated THP-1 cells were co-cultured with si-LATS1 BUC-87 cells culture supernatant for 72h. Macrophage polarization wasthen assessed by measuring the expression of several classical M1 markers (CD68+CD11b+CD86+) and M2 markers (CD68+CD11b+CD163+).
In the absence of si-LATS1 BUC-87 cell supernatant, our flow cytometric analysis showed a low expression of the M1 marker CD86 in and the M2 marker CD163. More importantly, in the presence of si-LATS1 BUC-87cell supernatant, THP-1 macrophages exhibited an M1 phenotype, represented by an increase in the surface expression of CD86(Figure 6B and 6C). These data suggested that the pryoptosisinduced bysi-LATS1was involved in M1 polarization of macrophages. To further confirm these results, the THP-1cells weredirectly co-cultured withBiu-87 cells transfected with LATS1 siRNA.Consistently,after 48h,the levels of CD86 were also upregulated in the THP-1cellswhen compared with those co-cultured withthe NC-transfected Biu-87 cells(Figure 6D and 6E). These results proved that si-LATS1 in BLCA cells could promote M1 polarization of THP-1 derived macrophagesin-vitro.
{kind=link}
{kind=link}