Development of four synthetic hexaploid wheat lines
Four F1 hybrid combinations were obtained from crosses between solid-stemmed Ma as the female parent and four different Ae. tauschii accessions AS78, AS92, AS95, and AS96 as the male parents in 2018. The true F1 hybrids were found cytologically to have a chromosome number of 21 (Fig. 1). The plant height and spike length of the F1 hybrids were similar to those of their female parent, but the number of tillers reached 10 to 20, which was similar to that of their male parent. The selfed seed set rates of F1 hybrid combinations were 16.71%, 16.48%, 20.36% and 25.77% for Ma/AS78, Ma/AS92, Ma/AS95, and Ma/AS96, respectively (Table 1). Then, four newly synthetic hexaploid wheat lines were developed from natural chromosome doubling of these true F1 hybrids, coded by Syn-SAU-116, Syn-SAU-117, Syn-SAU-118, and Syn-SAU-119.
Agronomic traits of the four synthetic hexaploid wheat lines grown in the field
The agronomic traits of the four synthetic hexaploid wheat varieties and their parents were evaluated in the field (Fig. 2; Table 2). The plant heights of all four synthetic wheat plants were higher than those of their parents (Fig. 2a), and there were very significant differences from their male parents. The plant heights of Syn-SAU-117 and Syn-SAU-119 were significantly different from those of their female parents. There was a very significant difference between Syn-SAU-116 and its female parent in the number of tillers. The spike lengths of the four synthetic wheat plants were longer than those of their parents, and there were very significant differences from those of their parents (Fig. 2b). The tiller number was between those of the parents. The seed length and width were similar to those of Ma but not as full as Ma (Fig. 2c). The synthetic wheat lines Syn-SAU-116, Syn-SAU-118 and Syn-SAU-119 had a higher self-seed setting rate, and Syn-SAU-117 had a lower self-seed setting rate. At the adult stage, all four newly synthetic hexaploid wheat lines were resistant (IT, 2-3) (Fig. 3), Ma was highly resistant (IT, 1) (Fig. 3), and all four Ae. tauschii accessions AS78, AS92, AS95, and AS96 were susceptible (IT, 7-8) according to Ref.50.
Observation of the stem solidity of synthetic hexaploid wheat lines grown in the greenhouse and field
In the greenhouse, the second internode marrow cavity of the stem base of the durum wheat Ma was filled with pith and was considered a solid stem with grade 5.0 (Fig. 4a; Table 2), while that of the common wheat CS had no pith, indicating a hollow stem with grade 1.0 (Fig. 4b; Table 2). The stem solidity of the second internode at the stem base of the four synthetic hexaploid wheat plants was not completely the same. Compared with the common wheat CS, Syn-SAU-116 and Syn-SAU-117 had a marrow cavity and stem wall between the second internodes at the base and were obviously thicker, being considered semisolid stems with grades 4.2 and 3.2, respectively (Fig. 4c, Fig. 4d; Table 2), while Syn-SAU-118 and Syn-SAU-119 were filled with pith in the second internode medullary cavity at the base, exhibiting solid stems with grade 5.0 (Fig. 4e, Fig. 4f; Table 2).
In the field, the marrow cavity at the base of the second internode of the durum wheat Ma was also filled with pith, indicating a solid stem with grade 5.0 (Fig. 5a; Table 2), while that of the common wheat CS had no pith, indicating a hollow stem with grade 1.0 (Fig. 5b; Table 2). The stem solidity of the second internode at the base of the four synthetic wheat lines was not completely the same. There was a very small marrow cavity between the second node at the base of Syn-SAU-117 and Syn-SAU-118, indicating semisolid stems with grades 4.1 and 4.5, respectively (Fig. 5d, Fig. 5e), and the stem wall is obviously thicker than that of CS. The medullary cavity of the second intersegment at the base of Syn-SAU-116 and Syn-SAU-119 was filled with pith, indicating a solid stem with grade 5.0 (Fig. 5c, Fig. 5f).
Therefore, the second internode at the base of the stem of Syn-SAU-117 was semisolid with a grade 3.2-4.1, while that of Syn-SAU-119 was solid with grade of 5.0 in both the greenhouse and field.
Determination of the lodging resistance of synthetic hexaploid wheat grown in the field
The breaking resistance of the second internodes of the stem bases of all four synthetic wheat plants was weaker than that of Ma, and the bending moment was less than that of Ma (Table 3). Compared with Ma, the lodging indices of both Syn-SAU-116 and Syn-SAU-119 were smaller, while the lodging indices of Syn-SAU-117 and Syn-SAU-118 were slightly larger (Table 3). The bending moment and lodging index of the second internodes of the stem bases of the four synthetic wheat plants were lower than those of the CS plants (Table 3). The breaking resistance of the second internode at the base of the stem of Syn-SAU-116 was less than that of CS. The breaking resistance of the second internode at the base of Syn-SAU-117, Syn-SAU-118 and Syn-SAU-119 was greater than that of CS. Syn-SAU-116 had the smallest lodging index, followed by Syn-SAU-119 (Table 3). Lodging resistance was expressed by the lodging index. The smaller the lodging index, the more resistant the plant was. The breaking resistance of synthetic hexaploid wheat was negatively correlated with the lodging index, and the bending moment had a very significant positive correlation with the lodging index (Table 4). Therefore, the synthetic hexaploid wheat Syn-SAU-116 had the strongest lodging resistance, followed by Syn-SAU-119.
The anatomical structure of stalks of synthetic hexaploid wheat grown in the field
The outer diameter of the culm of Ma was the largest (Fig. 6a; Table 5). The outer diameters of culms of the Ae. tauschii accessions AS78, AS92, AS95, and AS96 were significantly smaller than that of Ma, (Fig. 6c, Fig. 6e, Fig. 6 g and Fig. 6i; Table 5). The outer diameters of culms of the four synthetic wheat lines were smaller than that of Ma. Among them, Syn-SAU-116 had the largest outer culm diameter at 4209.18 μm (Fig. 6b; Table 5). However, the width of the pith cavity of Syn-SAU-116 and Syn-SAU-119 was 0 (Table 5), and the pith was full (Fig. 6b, Fig. 6 h), the same as that of Ma. Syn-SAU-117 and Syn-SAU-118 had very small widths of pith cavities (Fig. 6d, Fig. 6f), different from that of Ma. Syn-SAU-116 and Syn-SAU-119 had the largest ratio of wall thickness to outer culm diameter, reaching 50%, the same as that of Ma (Table 5). Syn-SAU-116, Syn-SAU-118 and Syn-SAU-119 had a larger percentage of mechanical tissue than Ma. Among them, Syn-SAU-119 had the largest percentage of mechanical tissue with 34.29%, and Syn-SAU-117 had a slightly smaller percentage of mechanical tissue than Ma with 21.48% (Table 5). Ma had a large number of vascular bundles, as many as 64, while that of four Ae. tauschii accessions was 27~40, much less than that of Ma (Table 5). The number of vascular bundles of the four synthetic wheat plants was 52~62.5, less than that of Ma (Table 5), but much larger than that of the corresponding male parents, which were Ae. tauschii.
In this study, it was indicated that the width of the pith cavity had a very significant positive correlation with the lodging index for synthetic hexaploid wheat (Table 6). The percentage of mechanical tissue had a negative correlation with the lodging index. The outer diameter of the culm of the second internode at the base of the stem had a significant negative correlation with the lodging index. The ratio of wall thickness to the outer culm diameter had a very significant negative correlation with the lodging index. There was no correlation between the number of vascular bundles and the lodging index.
Chromosomal observations of four synthetic hexaploid wheat lines
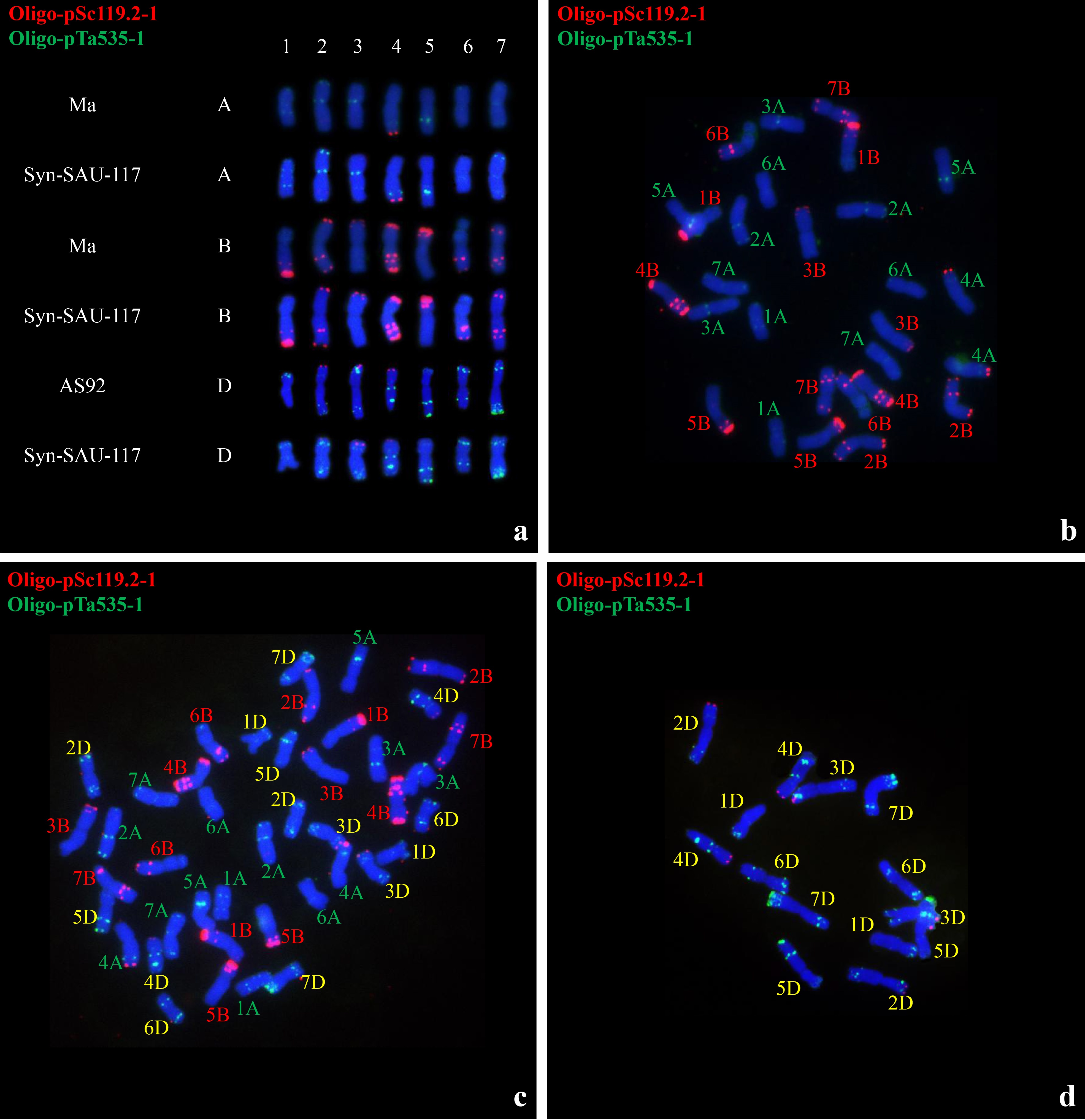
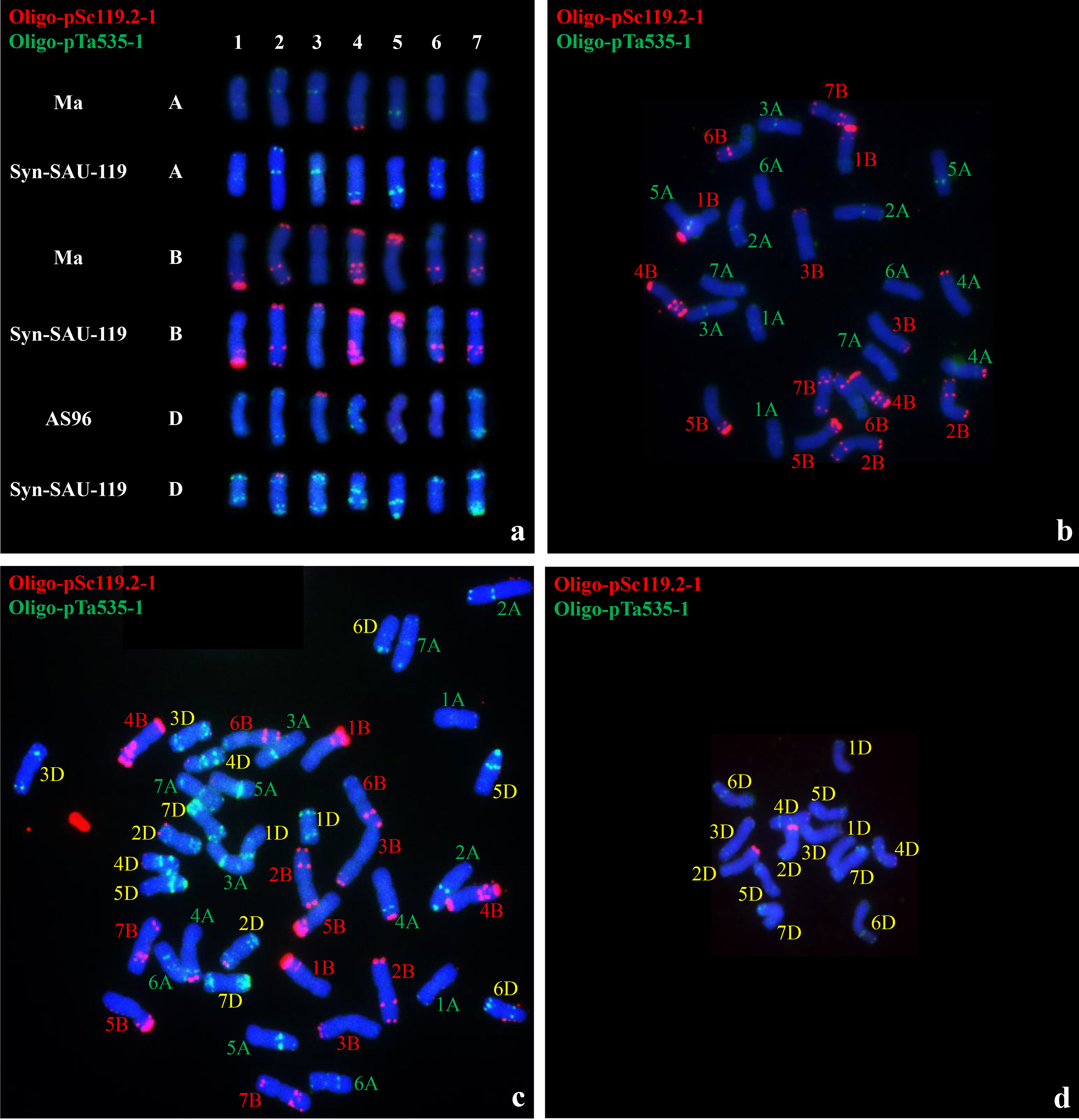
Analysis of root tip chromosome numbers showed that of 47 plants from four synthetic hexaploid wheat lines, 32 had 42 chromosomes, while 15 had 41 chromosomes (Table 7). Multicolor FISH was performed on the plants of four synthetic hexaploid wheat lines with 42 chromosomes using probes Oligo-pTa535-1 and Oligo-pSc119.2-1 (Fig. 7, Supplementary Information). The A-, B-, and D-genome chromosomes were distinguished according to Tang et al. (2014). The green-labeled Oligo-pTa535-1 probe mainly hybridized to the A- and D-genome chromosomes (Fig. 7, Supplementary Information). The red-labeled Oligo-pSc119.2-1 probe mainly hybridized to the B-genome chromosome, along with the signals at the end of the long arm of 4A and the end of the short arm of 2D, 3D and 4D (Fig. 7, Supplementary Information). Plants with 42 chromosomes were selected for the observation of chromosome pairing of PMCs in meiotic metaphase I. Most of the 42 chromosomes paired as bivalents (Fig. 8; Table 7), while a low number of univalent PMCs were also observed, indicating relative cytological stability.
{kind=link}
{kind=link}
{kind=link}