Metal ions removal efficiency
The toleration and adsorption to different metal ions of strain JC1 showed great difference. For instance, during S. rhizophila JC1 was respectively cultured in LB+Cu2+, LB+Cr6+, LB+Zn2+ that the concentration of metal ions was 40~120 mg/L , 40~200 mg/L, 40 mg/L for 24 hours, there were no significant difference in OD600 value. However, during the concentration of Zn2+ and Pb2+ were increased to 160 mg/L, the strain JC1 hardly grew. The adsorption efficiency was 79.8% when the concentration of Cu2+ was 120 mg/L-1, while it reduced to 26.1% when the concentration reached to 160 mg/L. More interesting was that with the concentration was 40~200 mg·L-1 of Cr6+ , there had no significant suppression on the growth of bacteria, but only showed maximum adsorption efficiency of 89.3% when the concentration was 80 mg/L. In terms of Pb2+ and Zn2+, the strain JC1 hardly showed adsorption ability (Fig. 1). Hence, this phenomenon indicated that the adsorption of metal ions by bacteria was selective, which was also suggested by Ye (Ye et al.2014)
Analysis of cells structure under metal stress by SEM
In this study, the morphological feature of strain JC1 comparison between metal ions stressed-cells and unstressed-cells was performed with SEM analysis. In different metal ions stress condition, different cell morphology was observed (Fig. 2). For unstressed-cells, the cell structure was intact, showed rod-shaped (Fig. 2a). However, the rod-shaped of JC1 after Cr6+ stress was almost impossible observed, there were crowds of particles gathered on the surface and the cells were wrapped in the membrane (Fig. 2b). In the case of Cu2+, cells showed shorter and brighter (Fig. 2c). The change caused by Pb2+ and Zn2+ was similar. In brief, the surface of the JC1 became rougher, but the difference was that the cells condensed into clusters after Zn2+ stress (Fig. 4d&e).
The change of cell morphology was the result of bacteria response to heavy metal stress. The decrease in cell size may be due to dehydration of the cells under Cu2+ stressed and also may be explained as a negative response of bacteria against further uptake of metal by decreasing the area of contact with the Cu2+( Mohite et al. 2018). The rough of cell surface might be interpreted as a strategy to accumulate more metal ions on the surface of cells as in case Pb2+ and Zn2+. For Cr6+, we speculated that due to the change of membrane permeability, a large amount of particles, formed through extracellular polysaccharides (EPS) binding with Cr6+, attached to the cell surface.
Analysis of chemical structure under metal stress by FT-IR
Studies had confirmed that the adsorption of heavy metals by bacteria is energy independent and can be mediated by functional groups (Zhang et al. 2017). In this study, the functional groups of strain JC1 for adsorption with four heavy metals was analyzed by Fourier transform infrared spectroscopy (FI-IR). Obviously, among four metal-laden samples, significant shifts of C-O-C and C=O were observed after the treatment of Cr6+, significant shifts of O-H was observed after the treatment of Cu2+ and Zn2+, significant shifts of C-O was observed after the treatment of Pb2+(Fig. 2f). According to the FI-IR analysis, we suggested that the binding by EPS was one of the important way for heavy metal detoxification. In addition, the detoxification that depends on the functional groups of the bacteria and the valence state of metal ions is selective (Ye et al. 2014, Zhang et al. 2017).
Analysis of element composition under metal stress by EDS
Generally, non-metallic elements such O, C, Si and non-toxic metal elements such as Ca2+, Fe2+/Fe3+ were the basic elements to maintain normal growth and metabolism of microorganism. However, some toxic metals such as Cr6+ or Pb2+ will exist in the cells through sites replacement due to atomic radius, cell structure. The result of element composition analysis after metal ions stress was shown in Fig. 3. Obviously, the corresponding metal elements were increased in the bacteria when they were stressed by different metals. Moreover, the metal content is consistent with the adsorption efficiency. Further analysis we found that, Cr6+ and Cu2+ mainly replaced the O element, while the Pb2+ and Zn2+ mainly replaced the C element, this may be related to the radius of the metal.
Analysis of physiological response of S. rhizophila JC1 under different metal ions stress
Determination of cell membrane permeability
The change of OM permeability under different heavy metal stress was shown in Fig. 4. Distinctly, the OM permeability of strain JC1 was significantly affected by the type and concentration of metal ions. What is more, its variation trend was positively correlated with the adsorption capacity of Cr6+ and Cu2+ (Fig. 1&4). However, it was interesting to note that the change induced by Pb2+ and Zn2+ were higher than Cr6+ and Cu2+, but the cell survival rate and adsorption capacity were contrary to this. Secondly, the higher the concentration of Pb2+ and Zn2+, the higher the change of OM permeability, but the cell survival rate and adsorption capacity were also contrary to this(Fig. 1&4). We speculated that: 1): detoxification factors for Cr6+ and Cu2+ were more abundant compared with Pb2+ and Zn2+ in cells of strain JC1, 2): cells will die when the membrane permeability changes exceeds its self-repair ability.
Similar to the OM permeability, the change of IM permeability were also affected by the type and concentration of metal ions. But it did not mean that the higher of the concentration of metal ions, the greater the change of membrane permeability, nor the stronger of the adsorption capacity. In summary, the change of membrane permeability was the key factor for bacteria to adsorb heavy metals, but when degree of change exceeded its self-repair capacity, it became non-resistant or low-resistant.
Analysis of changes in bioactive macromolecules
The excessive release of macromolecules such as proteins (Pr), phospholipid (PL), alkaline phosphatase (ALP) will definitely affect the normal function of cells. Without a doubt, with the change of membrane permeability, macromolecules will release to the outside of the cells. In terms of proteins and phospholipid, the variation trend was consistent with the change of membrane permeability (Fig. 4&5), which was also the reason that why cells hardly grow under the stress of Pb2+ and Zn2+. On the contrary, the release trend of alkaline phosphatase was consistent with the heavy metal adsorption capacity of strain JC1.We speculated that alkaline phosphatase participates in the hydrolysis reaction of microorganisms and generates phosphate ions (-PO43-) and free hydroxyl groups(-OH), then the -PO43- and -OH complexed with the positive metal ions such as Cr6+ and Cu2+, also with Pb2+ and Zn2+.
Transport proteome of S. rhizophila JC1
In total, 323 transport proteins belonging to nine major transport protein classes were predicted in the genome of S. rhizophila JC1 (4.28 Mb) (Table S1). Most of them were primary active transporters (TC#3), electrochemical potential-driven transporters (TC#2) and channels/pores (TC#1). They represented only 9% of the total number of predicted proteins. In addition, only 33%~50% of superior heavy metal-resistant bacteria which genome larger than 4 Mb(Rozycki and Nies 2009). The distribution of transport proteins was similar to these bacteria. Thus, the adsorption and resistance of S. rhizophila JC1 to heavy metals may be independent of the number of transporters, the types of transporter may be the key factor.
Heavy-metal transport proteins
Because cells do not contain the NADPH level required for reductase activities and there is a lack of methylation or other covalent modification mechanisms, some divalent metal ions cannot be discharged from cells independently. However, the three efflux systems (CDF, RND and P-type ATPases), containing almost all of the metal transport proteins, actively mediate the intracellular to extracellular excretion of divalent metal ions (Rozycki and Nies 2009). Therefore, the numbers and functions of transport proteins involved in the three efflux systems were analyzed in S. rhizophila JC1.
The cation diffusion facilitator (CDF) system
The CDF system members are chemiosmotic ion/proton exchangers that are involved in the efflux of divalent metal cations (Nies 2003) and the transport of metals from the cytoplasm across the cytoplasmic membrane into the periplasmic space (Higuchi et al. 2009) As shown in Table 1, two predicted proteins constituted the CDF system in S. rhizophila JC1. The czcD/zitB (JC1_GM001116) was predicted as Co2+/Zn2+/Cd2+ efflux system protein that included cation efflux, cation transporter ATPase C terminal as well as zinc transporter dimerization domains. Furthermore, its homolog was confirmed to bind Cu2+ and Ni2+ in Ralstonia metallidurans (Anton et al. 2004). The other CDF system member (TC#2.A.4.4.7) (JC1_GM001883) was predicted to be an integral membrane protein, which was confirmed as a lead (Pb2+) efflux transporter (PbtF) in Achromobacter xylosoxidans (Hložková et al. 2013), and it may provide sites for the targeted utilization of proteins. The sequences of these two proteins were provided in Additional file 1.
The resistance-nodulation-division (RND) efflux system
On the basis of transport substrate by bacteria, the RND was subdivided into six sub-groups: RND1 (Zn2+, Co2+ and Cd2+), RND2 (Co2+ and Ni 2+), RND3a (divalent cations), RND3b (monovalent cations), RND4 (Cu+ /Ag+) and RND5 (Ni2+)(Nies et al. 2003). Generally, these proteins are trimeric proteins that generate protein complexes named out membrane factor (OMF)(Koronakis et al. 2000) and a periplasmic adapter protein of membrane fusion protein(MFP)family(Akama et al. 2004).
Six heavy-metal efflux proteins were predicted in S. rhizophila JC1 (Table 1). Among them, two types of czcCBA operons (TC#2.A.6.1.15 and TC#2.A.6.1.16) form a complete cobalt–zinc–cadmium resistance system homologous to that in Escherichia coli (Tam et al. 2019). czcA is the RND protein, having at least one transmembrane domain and a membrane-spanning helical or beta-stranded domain that embeds in the membrane. czcC is a member of the OMP family that forms a trimeric channel and a long helical barrel that allows the export of a variety of substrates in Gram-negative bacteria. In S. rhizophila JC1, the czcCBA operon may transport Cd2+ and Zn2+, as well as Co2+, because there is a gene downstream of czcA that encodes a Cd2+/Zn2+-exporting ATPase (Fig. 6).
No regulatory gene was predicted near the two types of czcCBA operon, which was in contrast to the cnr in genome of R. metallidurans CH34 (cnrYXHCBA), czc in genome of R. metallidurans and Alcaligenes eutrophus (czcCBADRS/czcNICBADRS) and ncc in genome of Achromobacter xilosoxidans 31A (nccYXHCBAN)( Grass et al. 2000, Grosse et al. 2004). The amino acid alignment showed these paralogs share very low overall identity levels, as follows (Figure 6): JC1_001887 and JC1_003695 (czcC, OMF), 23.37% identity, JC1_001888 and JC1_003694 (czcB, MFP), 19.71% identity, and JC1_001889 and JC1_003693 (RND protein), 47.45% identity. Moreover, two additional genes (JC1_001886 and JC1_001890) were contained in the czc locus. JC1_001891, located downstream of the czcA gene, encodes the Cd2+/Zn2+-exporting ATPase (ZntA) that provides energy for Cd2+, Zn2+ and Co2+ efflux through hydrolysis. The low similarity level of these proteins suggested that they have bifurcated considerably in the process of evolution, they may have obtained unique way in protecting strain JC1 from being damaged by heavy metals (Valencia et al. 2013). The Ni2+/Co2+-uptake system was not predicted in the genome of strain JC1, but fortunately the Mg2+/Co2+-uptake system was predicted. It may be that S. rhizophila JC1 decreased its Ni2+/Co2+-detoxification function, but to maintaining homeostasis through controlling the efflux of outer membrane (Rozycki and Nies 2009). This finding meant the stepwise evolutionary from the ancestor of S. rhizophila had already able to deal with heavy metals (Nies 2003, Rozycki and Nies 2009).
One possible Cu(I)/Ag(I)-exporting RND4 protein (TC#2.A.6.1.8) was predicted in S. rhizophila JC1. This protein, as part of the copper resistance operon cluster, was encoded by chromosomal DNA and induced by Cu2+ (Moraleda et al. 2010). According to Nies (2006) classification of HME-RND, no specific RND3a (Zn2+) or RND3b (Co2+) transporters were predicted in S. rhizophila JC1. We speculated that Zn2+ and Co2+ detoxification by strain JC1 may depend on the chelation of related anionic groups.
P-type ATPase
S.rhizophila JC1 contains a relatively low number (5) of predicted P-type ATPases (Table 1). After concerned, copA involved a polypeptide domain of approximately 50 amino acid residues with two cysteines. Owing to the presence of this domain that the copA may be related to the detoxification of Cu+/Ag+, but ineffective to divalent ions that in contrast to copB. However, in Bradyrhizobium liaoningense by Liang (2016), copA is responsible for resistance to Cu+, Zn2+ and Cd2+. Moreover, copA induced gold (Au2+) detoxification due to a cytoplasmic metal-binding protein (Pontel et al. 2007).
The ZntA(JC1_GM001891) predicted in S. rhizophila JC1 was the P5-type ATPase, having plasma membrane C-terminal and N-terminal auto inhibitory domains. As described in Cupriavidus metallidurans, ZntA was the downstream gene of czcCBA operon that encoded the Cd2+/Zn2+-exporting ATPase, and its side chains, containing Met254, Cys476 and His807 contributed to Cd2+, Co2+ and Zn2+ binding and transporting, respectively(Smith et al. 2017).
Mercury detoxification
Three mercury detoxification proteins, MerP/T/R (JC1_GM001926–JC1_GM001928), were predicted in S. rhizophila JC1. MerP contributed to binding the highly toxic Hg2+, MerT was responsible for importing Hg2+ into cytoplasm, MerR was the regulator of mer operon (Silver and Phung 1996). Unlike in C. metallidurans CH34 (Rozycki and Nies 2009), there were no MerA and merRPT fragments predicted in S. rhizophila JC1. However, two copper efflux regulators (JC1_GM002046 and JC1_GM002150) and a redox-sensitive transcriptional activator SoxR (JC1_GM000920) were predicted.
In summary, the efflux system of S. rhizophila JC1 contained the same types of metal transport proteins as many other metal-resistant bacteria. Its low number of transport proteins may be the result of evolution.
Comparative genomics analysis
Molecular evolutionary analysis
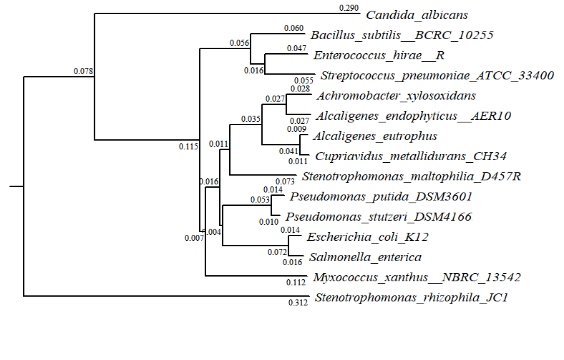
A molecular evolutionary analysis is an valuable measure to evaluate similarities or differences, and to match functional gene information, between model organisms and uncharacterized newly sequenced genomes(Gabaldón and Koonin 2013).
The evolutionary relationships of the 15 investigated bacteria were shown in Fig. S1. S.rhizophila JC1 had long-distance evolutionary relationships with other strains, which indicated that there may be great differences in gene expression levels. In addition, the lower number of transporters in genome of S. rhizophila JC1 compared with the other strains was the result of evolution.
Numbers of paralogs and transporters in the compared bacteria
For more information about the origin of metal-resistance gene of S. rhizophila JC1, the paralogs of predictive gene products of 15 investigated bacteria were analyzed by BLAST comparisons. On the whole, the frequency level of paralogs among compared bacteria was approximately 8% (Table 2). The relatively closest relationship with Salmonella enterica P-stx-12, has 30% paralogs, indicating that most of its increased genome size may be due to gene replication events during the bacterial species formation process.
In terms of most investigated strains, the frequency of paralogs of transporters was close to the 7% (Table 2). High percentages of paralogous protein-encoding genes were located on plasmids in S. rhizophila JC1 (28%), unlike the plasmid in Enterococcus hirae NCTC12204 (5%). When only considered the plasmid encoding paralogs of transporters, 29% of the S. rhizophila JC1 proteins represented paralogs. Therefore, duplicating genes encoding transport proteins, relocating them to plasmids and altering the substrate range and their products’ expression patterns may have been important evolutionary processes in the evolution of S. rhizophila JC1.
Comparison of transition-metal transporters
A higher number of heavy metal-specific transport proteins were predicted of strain JC1 through comparing with 15 investigated strains, especially proteins belonging to three efflux system (Table 3). For instance, the number of P-Type ATPases that contribute to Cd2+/Zn2+ even to Cu+/Cu2+ was nearly twice of other compared strains. Regrettably, the number of zinc transporters and mercuric transporters were relatively lower. The Mg2+/Co2+-transport protein (TC#9.A.40.1.2, JC1_GM001315) that belong to the HlyC/CorC family was predicted in S. rhizophila JC1. We speculated that it may have great detoxification ability to divalent cations due to its approximately 440 amino acids with an N-terminal four TMS domain. Here, we further speculated that the metal resistance gene of S. rhizophila JC1appeared to have evolved by horizontal acquisition or duplication (Rozycki and Nies 2009). The detoxification ability to heavy metal was perfect, even though the number of detoxification system had decreased.
Validation of predicted genes by qPCR
The expression level of 4 genes that respectively represents three efflux systems were identified by qPCR. Each gene to be validated was successfully amplified and produced a single band. CzcD, czcB and ZntA were up regulated after treated with Cr6+, Pb2+ and Zn2+ as compared to the CK. Cu2+-exporting ATPase and ZntA were up regulated in after treated with Cu2+ as compared to the CK. Among them, CzcD, czcB and ZntA exhibited higher expression levels in S. rhizophila JC1 treated with Cr6+ compared to samples treated with Pb2+ and Zn2+. The expression level of each gene used for identification showed significantly difference by P-value evaluation. The expression trend of 4 selected genes was consistent with the metal ions-removal efficiency and predicted results, which further confirmed that transport proteins play important roles in process of heavy metal adsorption.
{kind=link}
{kind=link}