Phenotypic analysis
The rooting phenotype of blueberry green cuttings varied significantly between the control and IBA treatment (Fig. 1a-b). The rooting percentage and average root number per cutting were both significantly promoted by exogenously applied IBA (Fig. 1c-d). There was no significant difference in average root length between these two treatments (Fig. 1e), suggesting that exogenous auxin IBA could enhance adventitious root formation without influencing the average length.
Microstructure observation of adventitious root formation
To clearly and definitely observe the AR developmental process against the stem of blueberry green cuttings, the microstructure was observed. The results showed that callus tissues started to form at 7 days after cutting (DAC) in the IBA treatment (Fig. 2a), adventitious root primordium initiation (rpi) was produced at 14 DAC (Fig. 2b), and adventitious root primordium (rp) developed at 21 DAC (Fig. 2c), rp then developed to adventitious root (AR) (Fig. 2d) and outgrowth from the stem was observed at 35 DAC (Fig. 2e). This process suggests that blueberry AR formation was initiated from non-root pericycle cells as a type of auxin-induction.
Analysis of plant hormones in blueberry green cuttings during AR formation
Changes in plant hormones during blueberry AR formation were analysed. The level of IAA exhibited a significant increase at 21 DAC under the IBA treatment (Fig. 3a-b), which might indicate that the higher IAA contributes to initiating adventitious root primordium (rpi). No significant difference in GA3 content was observed in the green cuttings between the control and IBA treatment (Fig. 3c). The content of ABA and zeatin under the IBA treatment was kept at a lower level during the whole trial, and a decreasing tendency of ABA and zeatin was recorded in the control treatment and obtained a similar level relative to the IBA treatment after 21 DAC (Fig. 3d), suggesting that a lower level of ABA and zeatin may facilitate AR formation.
De novo sequencing, assembly, and gene annotation in the RNA-seq analysis of blueberry stem
The basal stem of the cuttings were sampled every 7 days after insertion, and ten samples (CK7, CK14, CK21, CK28, CK35, T7, T14, T21, T28 and T35) were subjected to total RNA extraction and RNA-seq analysis. Total 40.73 - 46.23 million (M) pairs of 150 bp raw reads were generated from RNA_sequencing for each library (Table 1). After the data filtering, 212 million clean reads remained, out of which the clean reads number for each library ranged from 41.27 to 44.25 M (Table 1). Finally, 672606 transcripts and 308719 unigenes were de novo assembled with an average length of 735.87 bp (N50=1082) and 617.30 bp (N50=844), respectively (Table 2). After Blastx searching with a cut-off E-value of 10-5 against five databases, and the total unigenes annotated by NR database, GO database, KEGG database, eggNOG database and Swiss-Prot database were 107111, 53859, 6696, 101729 and 89910, respectively (Table 3).
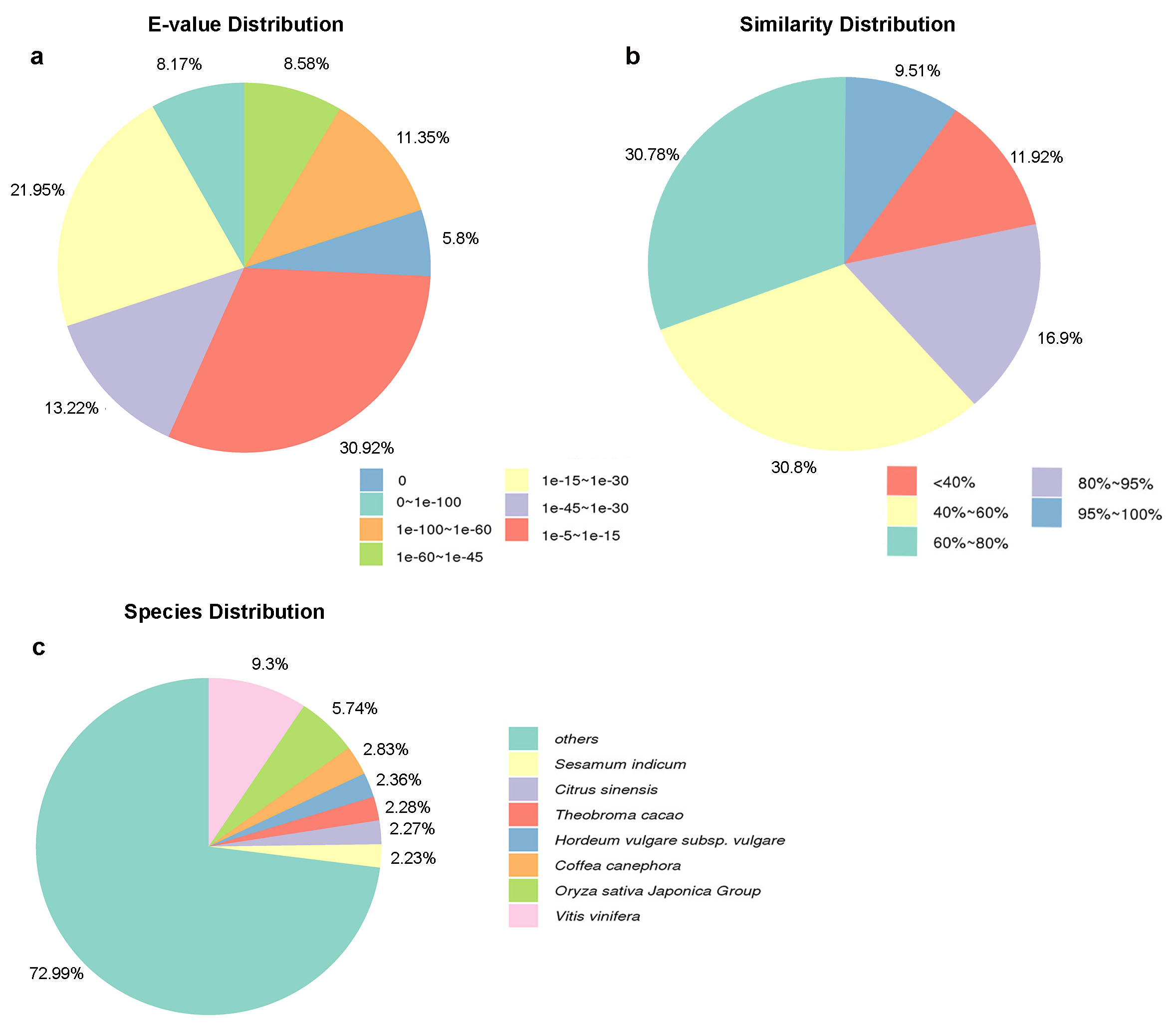
Based on NR annotations, 25.33% of the annotated sequences showed very strong homology (E-value < 10-60), 21.80% of the annotated sequences showed strong homology (10-60 < E-value < 10-30), and an additional 82.87% of the annotated sequences showed homology (10-30 < E-value < 10-5) to available plant sequences (Additional file1: Fig. S1a). The similarity distribution was comparable, with 26.41% of the sequences showing similarities higher than 80% and 73.5% of the hits showing similarities of 0-80% (Additional file1: Fig. S1b). With respect to species, 72.99% of the unique genes showed high matches with sequences from other species, including Vitis vinifera, Oryza sativa Japonica Group and Coffea canephora (Additional file1: Fig. S1c).
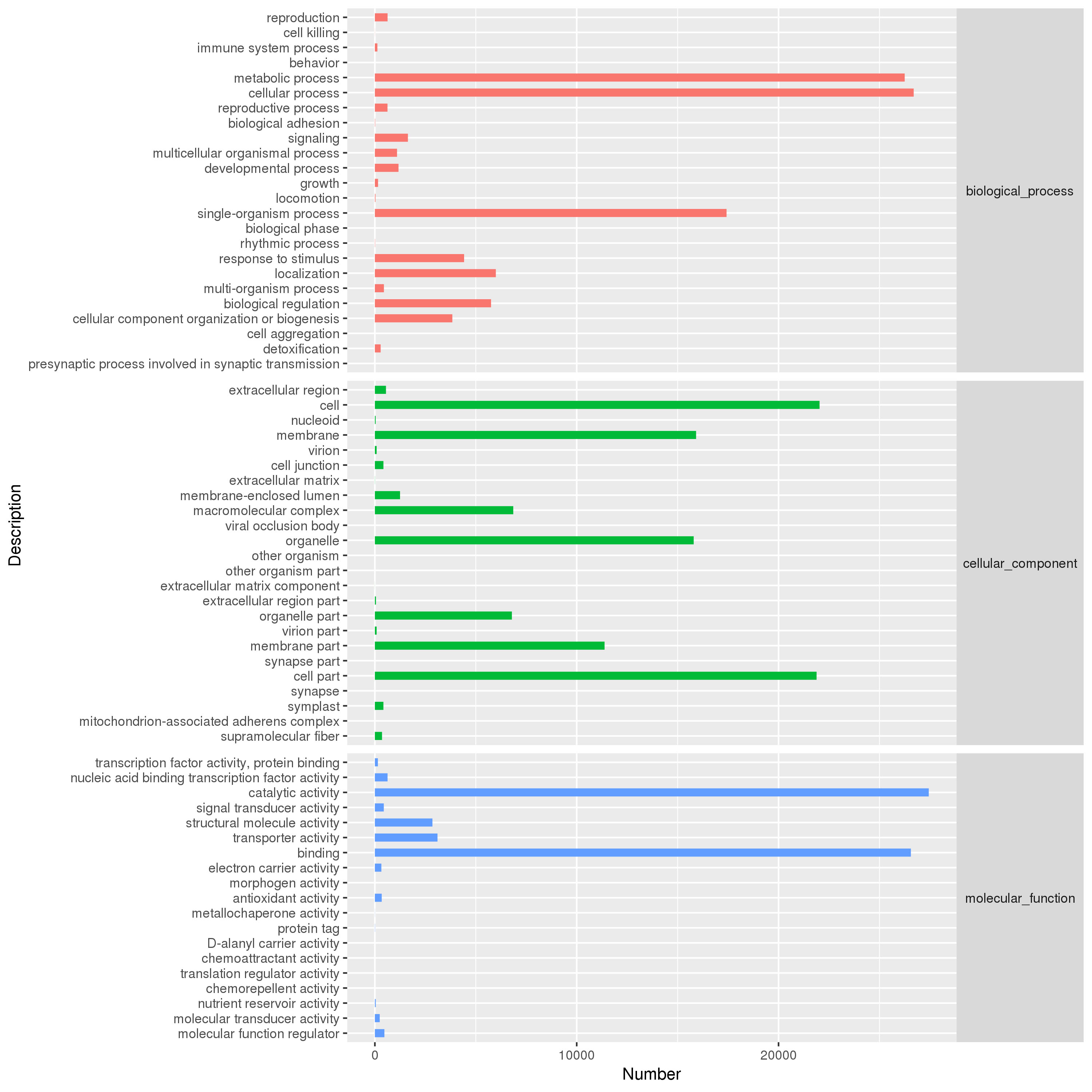
The GO analysis was performed with Blast2GO software. Out of 308719 unigenes, 263143 were classified into the “biological process”, “cellular component” and “molecular function” categories (Additional file2: Fig. S2). This classification provided some information on the percentage of blueberry unigenes in different signal transduction, catabolic and anabolic processes. For the biological process category, the majority of unigenes was grouped into “metabolic process”, “cellular process” and “single-organism process”, which accounted for approximately 72.83%. In the cellular component category, the unigenes were mainly distributed into “cell”, “cell part”, “membrane”, “membrane part” and “organelle”, accounting for approximately 83.72%. For the molecular function category, a large number of unigenes was distributed into “catalytic activity” and “binding”, which accounted for approximately 86.26% (Additional file2: Fig. S2).
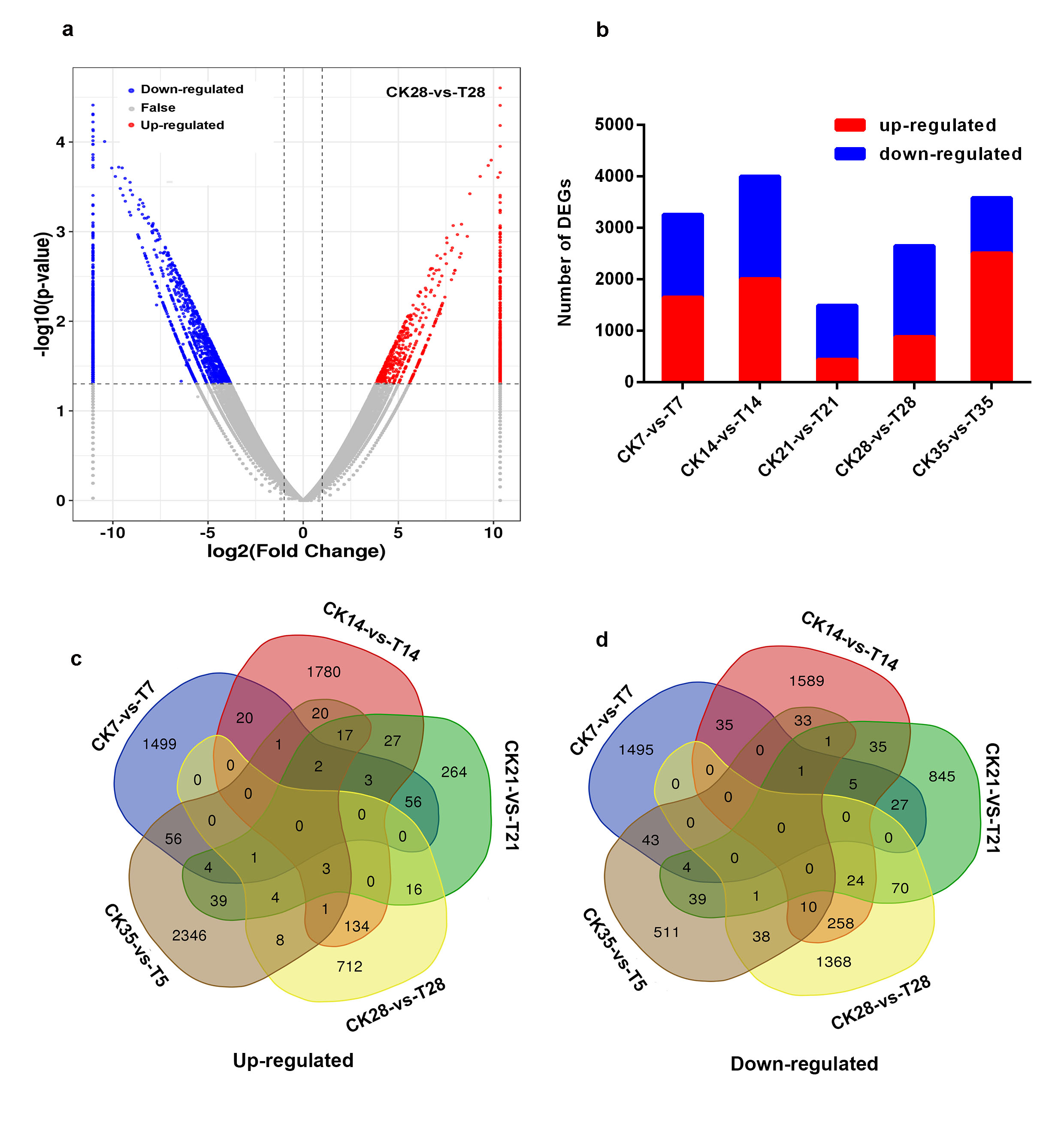
Based on RSEM quantitative software, the FPKM value of the unigenes was calculated between the control (CK) and IBA treatment (T). The DEGs were identified by pairwise comparisons of ten libraries, i.e., CK7 vs. T7, CK14 vs. T14, CK21 vs. T21, CK28 vs. T28, and CK35 vs. T35 (Additional file3: Fig. S3a). A total of 14970 DEGs were determined, out of which 7467 were upregulated and 7503 were downregulated (Additional file3: Fig. S3b). For each sampled period, 3252 DEGs were detected between CK7 and T7 libraries, and these unigenes were directly affected by IBA treatment and might be associated with callus tissue formation (Fig. 2a, Additional file3: Fig. S3b). In CK14 vs. T14, 3999 DEGs were identified, which might be related with rpi formation (Fig. 2b, Additional file3: Fig. S3b). In CK21 vs. T21, 1488 DEGs were identified, which contributed to rp formation (Fig. 2c, Additional file3: Fig. S3b). In CK28 vs. T28, 2648 DEGs were identified, which contributed to AR formation (Fig. 2d, Additional file3: Fig. S3b). And in CK35 vs. T35, 3583 DEGs were detected, which might have contributed to AR outgrowth and development (Fig. 2e, Additional file3: Fig. S3b). However, there were no commonly upregulated or downregulated DEGs at all sampled periods as illustrated in the Venn diagram (Additional file3: Fig. S3c-d), suggesting that DEGs might play special roles during AR formation, outgrowth and development.
DEGs enriched in the auxin signalling pathway
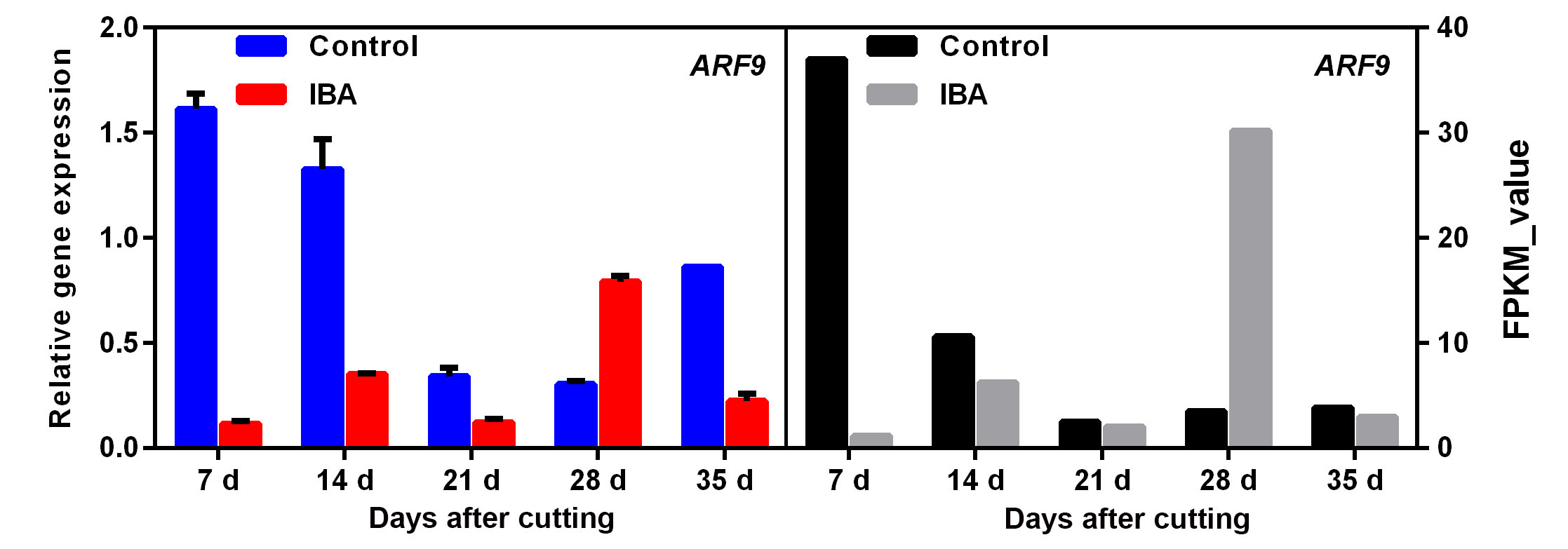
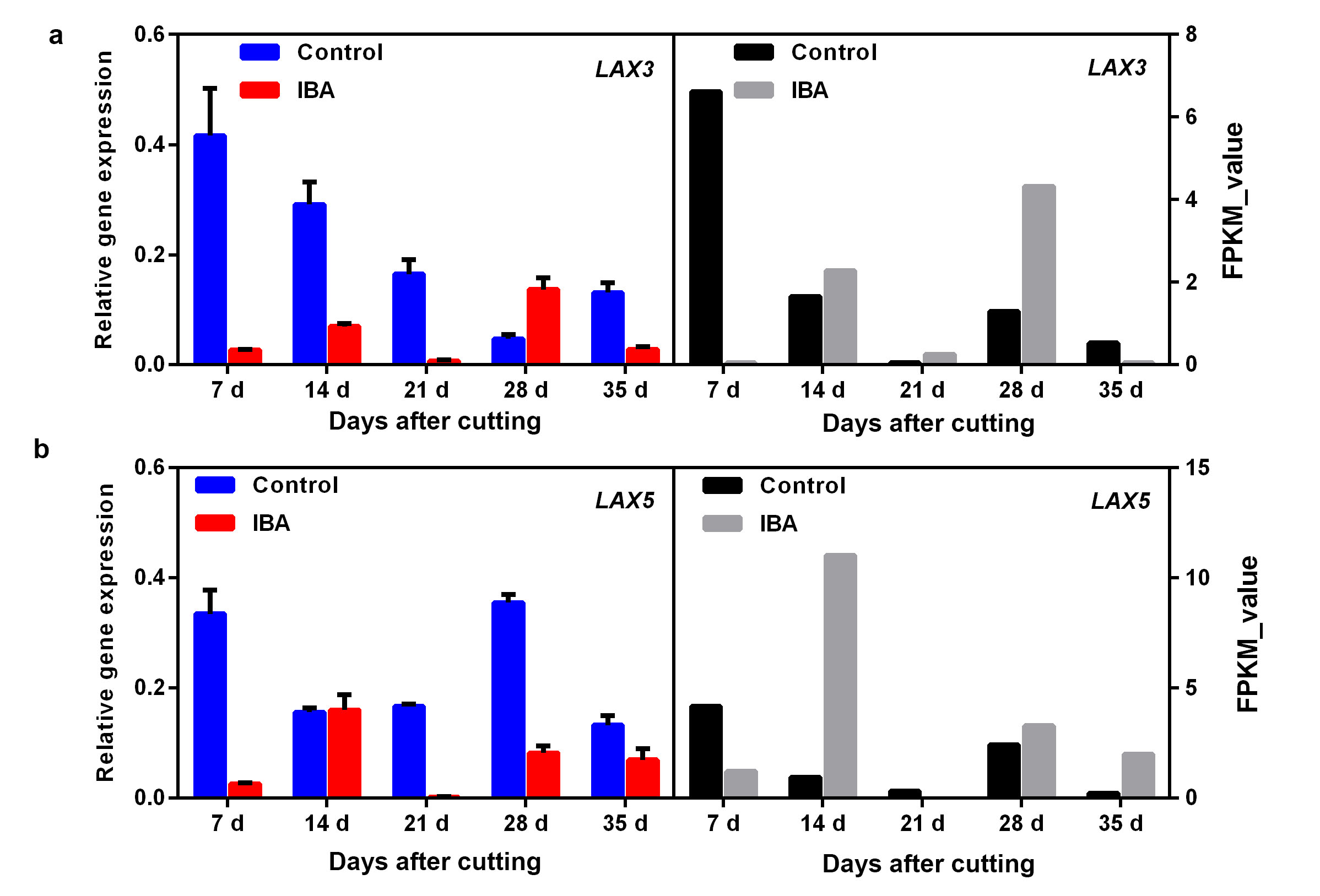
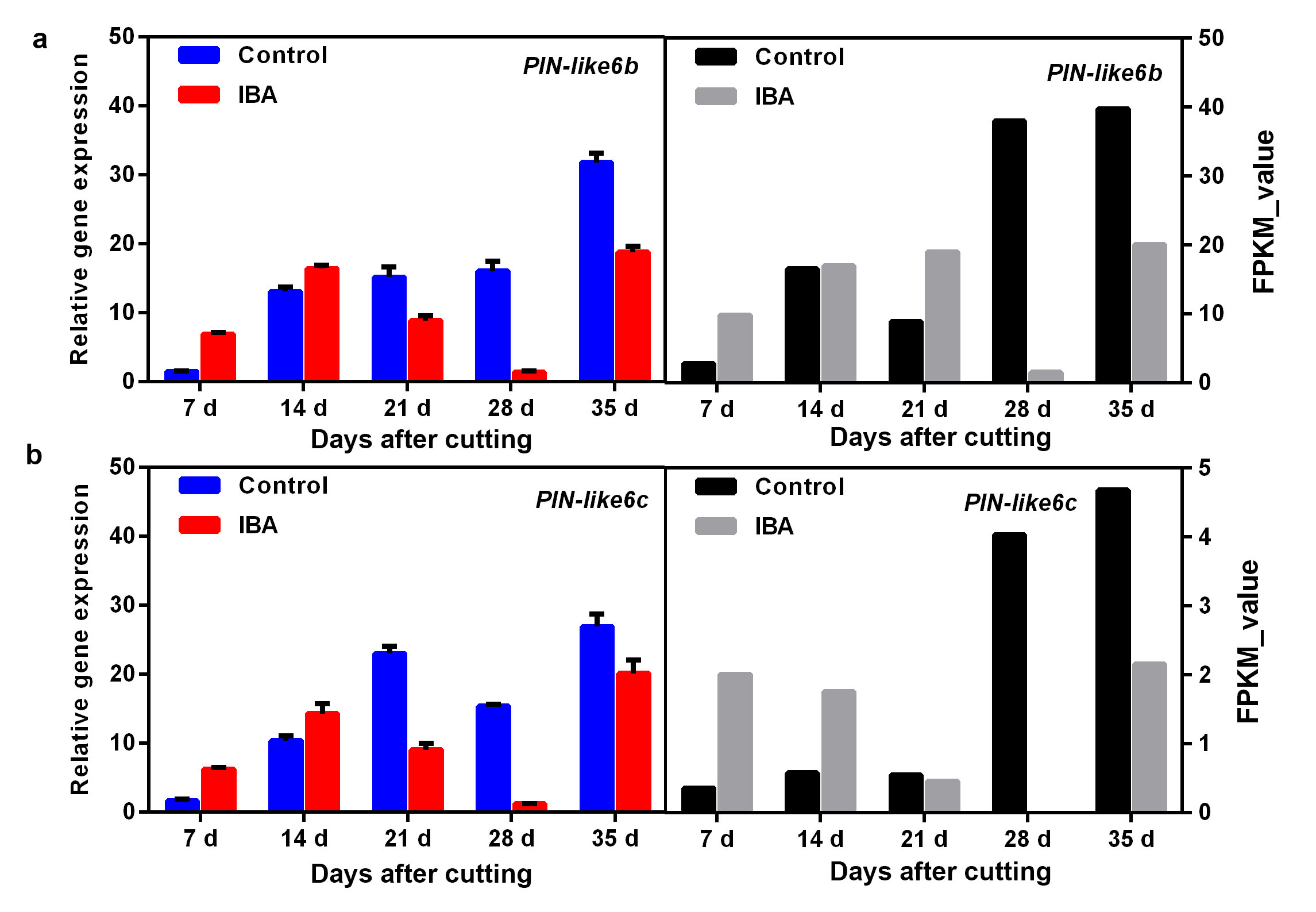
Auxin has been proven to play key roles in promoting AR formation; therefore, the DEGs in the auxin-signalling pathway were annotated and further analysed. A total of 29 auxin-related DEGS were mapped (Fig. 4). Of these DEGs, there were ten unigenes belonging to auxin-responsive proteins, including four ARFs and six SAURs. Six auxin transporter-like genes were also identified, including three influx carriers (AUX22, AUX-LIKE 3 (LAX3), and LAX5) and three efflux carriers (PIN-LIKE 6 (PIL6s)). All these DEGs were upregulated during AR development to some extent (Fig. 4). To verify the results of the comparative transcription analysis, two auxin responsive factor ARF genes (i.e., ARF7 and ARF9) and six auxin transporter genes (i.e., AUX22, LAX3, LAX5 and PIL6a-6c) were selected to identify the DEG expression profiles by qRT-PCR. RNA_seq showed that ARF7 and ARF9 exhibited a significant transcript peak at 28 DAC in the IBA treatment, while both these ARFs showed a decreasing trend in the control (Fig. 5a, Additional file4: Fig. S4). RNA_seq data revealed that AUX22 was kept at a lower expression in the control but showed a significant upregulation at 7 DAC – 21 DAC in the IBA treatment (Fig. 5b). Its homologous genes LAX3 and LAX5 showed a decreasing tendency in the control, which were significantly upregulated at 14 DAC and 28 DAC in the IBA treatment (Additional file5: Fig. S5). These results suggest that these genes might play an important role in auxin transport and contribute to auxin asymmetrical distribution, which was facilitated to induce the adventitious root primordium initiation at the early stage of AR formation. For the three auxin efflux carriers PIL6a-6c, in the control treatment, these three genes showed low expression at 7 DAC – 21 DAC and were upregulated until 28 DAC. However, after applying IBA, PIL6a was significantly upregulated at 14 DAC (Fig. 5c) and PIL6b and PIL6c showed a remarkable increase from 7 DAC and peaked at 35 DAC (Additional file6: Fig. S6), suggesting that auxin enhances the expression of PIL6s and thus promotes AR formation. qRT-PCR data indicated a similarity with transcript information.
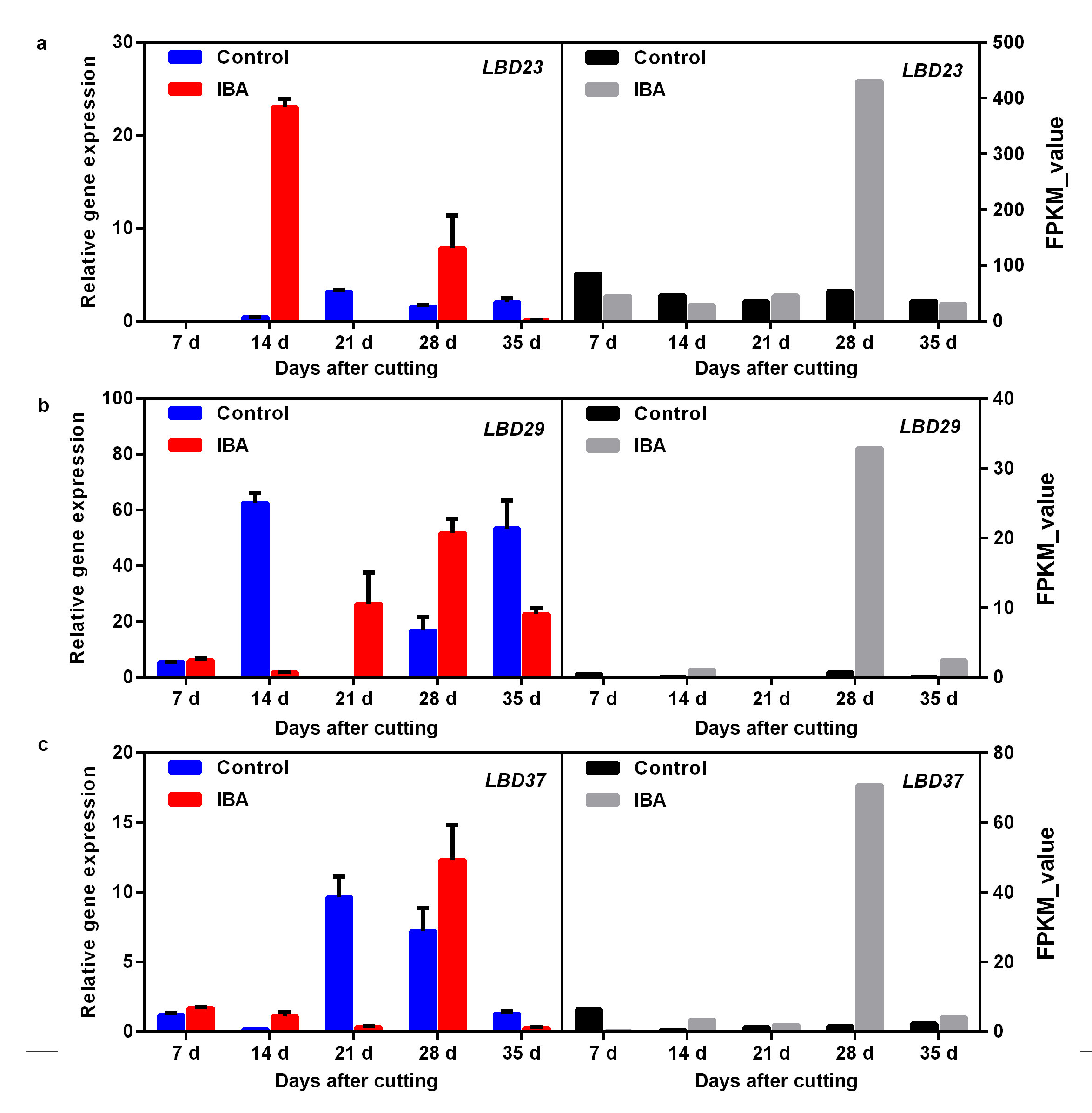
Transcription factor LATERAL ORGAN BOUNDARIES DOMAIN/LOB domain-containing proteins (LBDs) were suggested to regulate AR formation as downstream target genes of the ARF family. In the present work, thirteen homologue genes of LBDs were identified based on NR annotation (Fig. 4), and the RNA-seq data revealed that all these LBDs were up- or downregulated at different stages to a certain extent, especially at 28 DAC. Four out of these 13 LBDs (i.e., LBD16, 23, 29 and 37) were representatively selected to confirm their expression profiles by qRT-PCR, and the results indicated that LBD16 and LBD37 showed good reproducibility with the RNA-seq data. Moreover, the qRT-PCR indicated that LBD23 was upregulated again at 14 DAC in the IBA treatment and LBD29 showed continuous upregulation from 21 DAC to 35 DAC in the IBA treatment and was also upregulated at 14 DAC and 35 DAC in the control (Fig. 5d, Additional file7: Fig. S7).
DEGs involved in root primordium formation
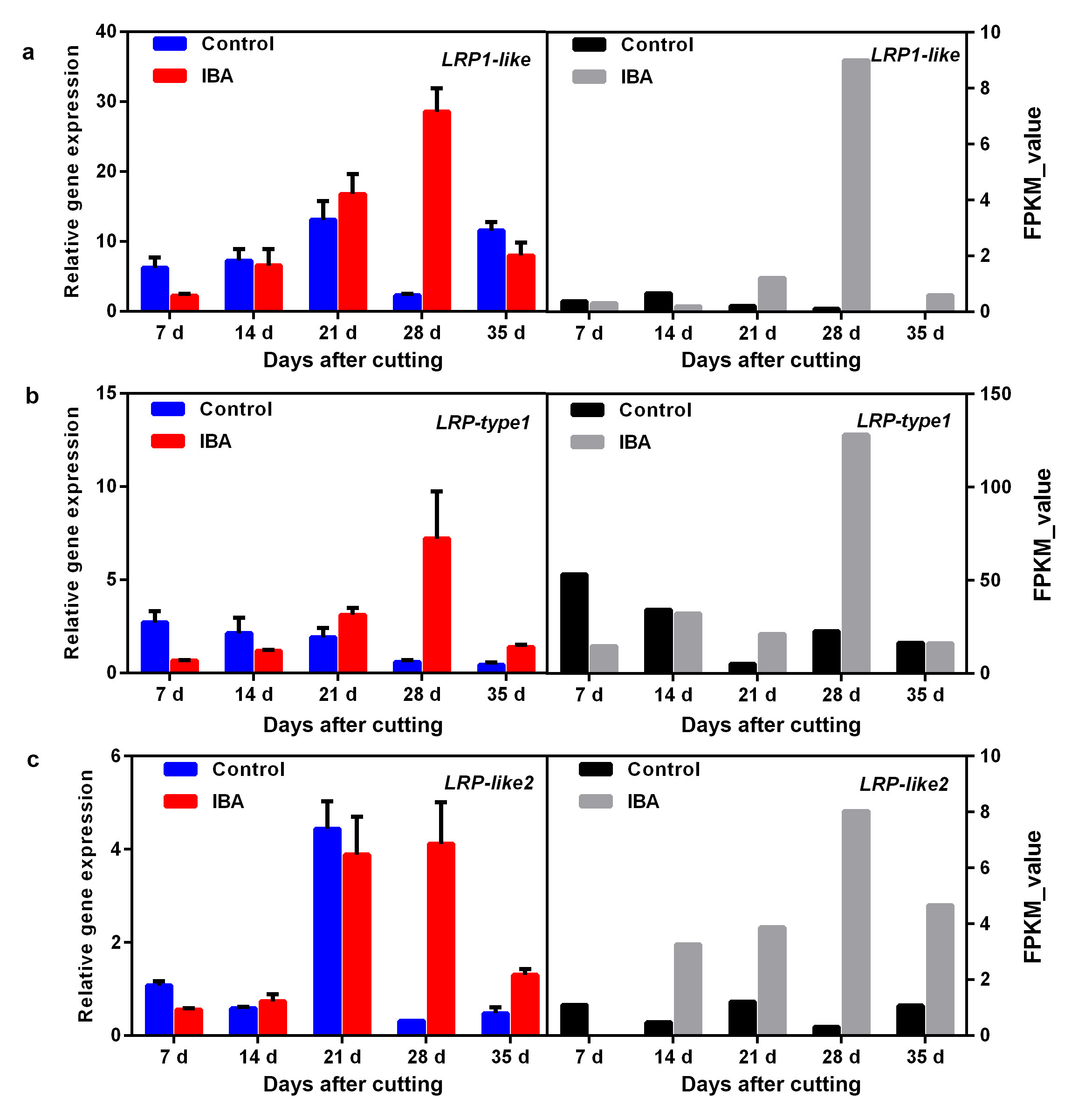
Based on the NR annotation, six rooting-related DEGs were obtained including four homologue genes of lateral root primordium 1 (LRP1), one putative root meristem growth factor 9 (RGF9), and one dormancy-associated protein homologue 3 (DRMH3) (Fig. 4). The RNA_seq data showed that LRP1 was significantly upregulated at 28 DAC (Fig. 5e), and its homologous genes LRP1-like, LRP-type1, LRP-like2 exhibited continuous upregulation after 14 DAC and peaked at 28 DAC in the IBA treatment (Additional file8: Fig. S8). Obvious upregulation of RGF9 was observed at 7 DAC, 14 DAC and 28 DAC (Fig. 5f), DRMH3 showed significant upregulation at 28 DAC in the IBA treatment (Fig. 5g). The qRT-PCR analysis of RGF9 indicated good consistency with the RNA_seq data, and the expression of DRMH3 in the IBA treatment showed good consistency with the RNA_seq data but was continuously expressed at a high level in the control treatment (Fig. 5g).
Putative gene regulatory networks that control blueberry AR formation
According to the known regulatory networks reported previously for root formation in Arabidopsis and other plant species, the regulatory pathway that controls blueberry AR formation was derived. It was speculated that IBA would induce the expression of auxin responsive factors ARF7/9 to perceive auxin signalling, whereas ARF7/9 directly or indirectly affected the downstream target LBDs genes to establish AR founder cells with nuclei migration. Then, auxin polar carriers, including influx carriers AUX22 or LAX3/5 and efflux carriers PIL6s, would be upregulated to facilitate the establishment of auxin asymmetric distribution, which includes AR primordium formation. Finally, the AR primordium transforms to the AR apical meristem and outgrowth from the cuttings under the effect of LRP1, RGF9, DRHM3 and other genes (Fig. 6).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}