Diversity of fish: A total of 21 species of catfishes from 15 genera and eight families were collected and checked for monogenoids (Table 1) (Fig. 1). Mystus and Glyptothorax had the highest diversity (three species each), followed by Batasio and Creteuchilolansis (two species each). The remaining genera had the least diversity (one species each). Bagridae was the most diverse family (six species), followed by Sisoridae (four species), Siluridae (three species), and Eristidae and Amblyceptidae (one species each).
Patterns of parasite diversity
The final host-parasite dataset included 25 parasite records from 21 host species from eight different families (Table 1). Of the 21 host species, 15 (62.5%) were infected with 11 previously described species from four genera (Bifurcohaptor, Cornudiscoides, Mizelleus, and Thaparocleidus) and 14 new species from four genera (Bychowskyella, Cornudiscoides, Gyrodactylus, and Thaparocleidus) (Fig. 2). The remaining six catfish species were not infected, likely due to the relatively small numbers (< 5) of specimens from these species were collected. These six species were Creteuchiloglanis kamengensis (Jayaram, 1966), Creteuchiloglanis sp., Pachypterus atherinoides (Bloch, 1794), Heteropneustes fossilis (Bloch, 1794), Nangra assamensis (Sen and Biswas, 1994), and Olyra longicaudata (McClelland, 1842).
The identified parasite species belonged to six known genera (Bifurcohaptor, Bychowskyella, Cornudiscoides, Gyrodactylus, Thaparocleidus, and Mizelleus), two families (Dactylogyridae and Gyrodactylidae), two orders (Dactylogyridea and Gyrodactylidea), and one subclass (Polyonchoinea) of the class Monogenoidea. Thaparocleidus was the most diverse genus (12 species; 48%), followed by Cornudiscoides (four species; 16 %), Bychowskyella and Gyrodactylus (three species each; 12 %), and Bifurcohaptor (two species; 8 %) and Mizelleus (one species; 4 %) (Fig. 3). Except for Cornudiscoides proximus and Bifurcohaptor indicus, which infected two hosts: Mystus dibrugarensis and Mystus tengara, other parasite species were restricted to one host (specialists). The infection levels ranged from 0 to 5 species of monogenoids (on Mystus dibrugarensis and Wallago attu). All species except Gyrodactylus sp. n. 3 (which infected the skin of A. arunachalensis) inhibited gills, confirming that the gills of fish species are the preferred biotope for Indian monogenoids.
In terms of heterogeneity, Bagridae was the most diverse family (four parasite genera), followed by Siluridae and Sisoridae (two parasite genera each), and Erithistidae and Amblyciptidae (one parasite genera each) (Fig. 4). Wallago attu and Mystus dibrugarensis had the highest numbers of monogenoid species (five parasite species each), followed by Pterocryptus indicus, Mystus bleekeri, Mystus tengara, and Ompok pabda (two parasite species each); the remaining nine hosts each had one monogenoid species (Fig. 2).
Only three Gyrodactylus species have been described from Indian catfishes: G. gusevi Dubey, Gupta and Agarwal 1990 and G. neonephrotus malmbergi Singh and Agrawal, 1994 both from Heteropneustes fossilis, and G. mizellei Dubey, Gupta and Agarwal 1990 from Mystus vittatus. Therefore, the record of three new Gyrodactylus species from catfishes of Arunachal Pradesh was a special finding (Table 1).
New host and geographic distribution records: Arunachal Pradesh represents a new geographical locality record for all 11 previously described parasite species (Table 1). This considerably expands their geographic distribution from North and South India to also include the country’s far extreme northeastern corner. These species cover a range of more than 2000 km. Two species of catfish represent new host records for previously known species of parasitic monogenoids—Ompok pabda (for Thaparocleidus malabaricus) and Mystus bleekeri (for Cornudiscoides agarwali) (Table 1). Ten species of catfish represent first host records for (new species of) parasitic monogenoids: Glyptothorax cavia, Glyptothorax mibangi, Glyptothorax dikrongensis, Pterocryptus indicus, Batasio batasio, Batasio fasciolatus, Gagata cenia, Mystus dibrugarensis, Pseudolaguvia viriosa, and Amblyceps arunachalensis (Table 1).
Interestingly, we found new species of monogenoid only in catfishes that can be found only in Arunachal Pradesh and/or other parts of Northeast India and, therefore, had never been sampled before. Conversely, we found new geographic records for—but no new species of—monogenoids when we screened fish that had been sampled before in other parts of the country. The lone exception to this pattern was Mystus bleekeri. This species had been studied for monogenoids and was known to harbour six monogenoid species: T. pusillus (Gusev, 1976) Lim, 1996), C. bleekerai Agrawal and Vishwakarma, 1996, C. geminus Gusev, 1976, C. gusevi Agrawal and Vishwakarma, 1996, C. susanae Agrawal and Vishwakarma 1996, and C. tukarami Agrawal and Vishwakarma, 1996 (see Pandey and Agarwal 2008). In the present study, we found two species from M. bleekeri (Cornodiscoides sp. n. 1 and Cornudiscoides agarwali Agrawal and Vishwakarma, 1996) but, surprisingly, found none of the six previously identified species mentioned above. Tripathi et al. (2019) reviewed this problem in detail and concluded that earlier authors probably confused M. vittatus with M. bleekeri. Examining the samples of M. vittatus and finding thereon worms described from M. bleekeri would confirm this assumption.
Impact of host habitat
In this study, all infected catfish were demersal except for Pterocryptis indicus and Glyptothorax cavia, which were benthopelagic (Table 1) (see Froese and Pauly, 2018). A comparative analysis of the parasite species’ richness across 15 catfish shows that both benthopelagic and demersal catfishes were infected with monogenoids. However, demersal fish were infected with 21 parasite species, while benthopelagic fish were infected with only three parasite species. This is likely due to sampling bias, as the demersal fish outnumbered the benthopelagic fish in our sample. So, in a broad sense, our results are inconsistent with previous studies; for example, Luque et al. (2004) reported that host depth range had no effect on ectoparasite species richness.
Impact of host body size
The size of the host body is an important ecological determinant of parasite species richness (Morand 2015). We observed heterogeneous results concerning the correlation between body size and parasite species richness. W. attu had the most monogenoid species (five), possibly due to its large size (total length of up to 240 cm) (Froese and Pauly 2018), which provides more available niches for parasitic colonisation (see Sasal et al. 1997). Alternative justifications might also explain this pattern—for example, since W. attu is a large potamodromous fish covering an extensive geographical range, it is exposed to more parasite species than other catfishes (see Morand 2000). The case of M. dibrugarensis, on the other hand, contradicts the notion that the larger the host size, the higher the parasite richness. M. dibrugarensis is a small silurid (total length of up to 9.50 cm) (Bailung and Biswas 2015), yet it harboured as many different species as W. attu (five) (Table 1).
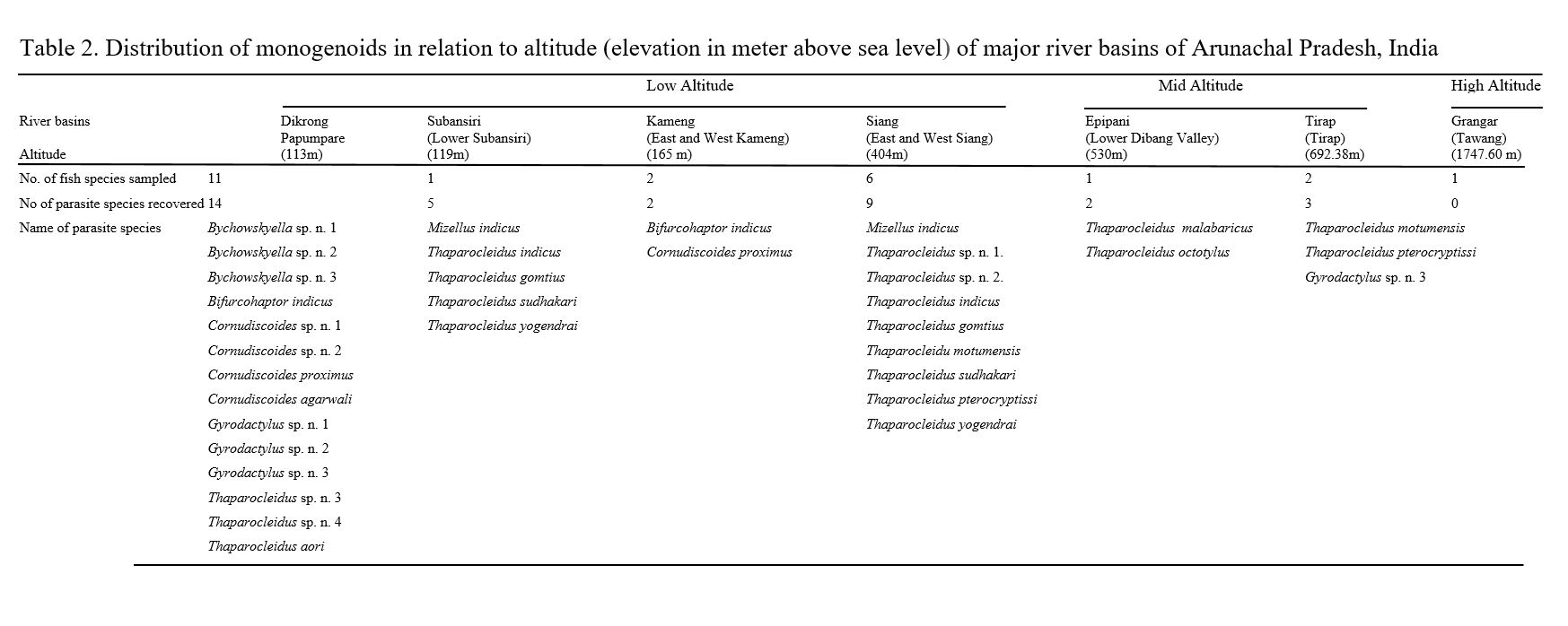
Impact of elevational gradient: A major issue in modern parasite ecology is the current lack of knowledge of the factors that influence the diversity and distribution of parasites—and, consequently, disease outbreaks (Hawkins et al. 2003). Unfortunately, the main drivers of parasite diversity are still poorly understood (Poulin and Morand 2004). The data, though preliminary, indicates a negative correlation between the species richness of parasitic monogenoids and the 1747.60-m elevational gradient (Table 2) (Fig. 5). Lower altitudes were home to 83.33% of the specific diversity of monogenoids, while mid altitudes were home to 16.66% of the specific diversity. At high altitudes, no parasites were found. Water temperature is one of the most important environmental variables influencing the distribution of monogenoids, with higher water temperatures increasing their reproduction rate and population growth (Jansen and Bakke 1991; Anderson and Buchmann 1998). We suggest that the observed general decrease in monogenoid species richness along an altitudinal gradient is caused by the corresponding decrease in water temperature because temperature is inversely related to elevation.
Host specificity of monogenoids
The rivers Ganges (in north India) and Brahmaputra (in northeast India) originate from glaciers on the southern and northern slopes of the Himalayan mountains, respectively. They take different paths before joining in the Bengal Basin and debouching to the Bay of Bengal (Subramanian and Ramanathan 1996, as cited in Kuehl et al., 2005). Then, how did ten monogenoid species previously described from the north India and south Indian river waters end up in the basins of the River Brahmaputra in Arunachal Pradesh even though these basins are not connected in India?
In general, the distribution of parasites follows the distribution of their hosts (Rohde 1993; Poulin 1998). This seems to hold especially true for monogenoids, which mandatorily depend on their fish hosts to complete their life cycle. Existing explanations for how fish (and their parasites) colonise unconnected water bodies involve either dispersal or vicariance—vicariance-based scenarios are generally favoured (see, for e.g., Berra 2001, Zanata et al. 2005). In either case, the presence of North and South Indian species of host fish and their monogenoids in Arunachal waters indicates that the host fish must have brought their parasites with them as they travelled from North or South Indian waters to the Brahmaputra River system of the Arunachal Pradesh region. Moreover, these worms did not engage in host switching (i.e., they remained on their original hosts and did not colonise new potential host species). Thus, our observation that previously non-sampled catfishes harboured new monogenoid species while previously sampled catfishes harboured already known monogenoid species supports the prediction of Whittington et al. (2000) that the monogenoids are probably the most host-specific of all fish parasites.
Variety of species on Wallago attu: Wallago attu presented a unique problem. This fish—sometimes known as the freshwater shark—is a commercially and recreationally important catfish native to Indian subcontinent (Froese and Pauly 2018). The species has been listed as endangered (Mijkherjee et al. 2002) and vulnerable (Ng et al. 2019) because of a rapid population declines throughout its range due to their over-exploitation for food, ornamental and recreational purposes, pollution, and environmental degradation, among other reasons. The systematics of the monogenoids of this host fish have become unmanageable due to new species descriptions over the last three decades. To date, approximately 40 nominal monogenoid species from 10 genera have been recorded from W. attu (Lim et al. 2001; Pandey and Agarwal 2008). Much of the information on these parasite taxa has been published in obscure, often inaccessible local journals, thus giving rise to numerous nomenclatural synonymies in the literature. Not surprisingly, our sampling efforts from W. attu in Arunachal Pradesh consistently yielded only five species: Thaparoleidus gomtius, Thaparocleidus yogendraii, Thaparocleidus indicus, Thaparoleidus sudhakari, and Mizelleus indicus. While we have no intention of discrediting the observations of previous authors who described these species, we also would not be surprised if many of the previously described species are spurious. In any case, the taxonomy of monogenoids from threatened W. attu is in a state of considerable confusion and instability. The parasites may have a negative impact on threatened hosts, in part because they can accelerate severe population declines (De Castro and Bolker 2005; Blehert et al. 2009) and, as a result, the extinction risk of their host species (Altizer 2007; Heard et al. 2013). Thus, a precise validation of the taxonomic assignments of monogenoids from W. attu is required, particularly based on critical evaluations and DNA-based taxonomy.
Missing parasite groups: So far, the following seven monogenoid genera have been identified from freshwater Indian Siluriformes: Ancylodiscoides (Yamaguti, 1937), Bifurcohaptor (Jain, 1958), Bychowskyella (Akhmerov, 1952), Cornudiscoides (Kulkarni, 1969), Mizelleus (Jain, 1957) Quadriacanthus (Paperna, 1961), and Thaparocleidus (Jain, 1952) (Pandey and Agarwal 2008). Of these species, we failed to find only two: Ancylodiscoides and Quadriacanthus. Since the present study sampled only 21 out of 69 Siluriformes, the absence of these two genera could simply be due to the small sample size.
Future of parasitological research in Arunachal Pradesh: Arunachal Pradesh has a relatively short history of modernisation, having begun only after India’s independence in 1947 (Planning Commission of India 2018). The state has just one university and no research centres or laboratories for studying parasitology. Consequently, the fish parasites inhabiting this area, including monogenoids, have remained largely unknown. In fact, in 2011, we were the first group of researchers to study fish parasites of this region. Two of our previous (unpublished) studies, which were conducted as parts of projects sponsored by the Government of India [UGC; 39–603/2010 (SR) and DST; SR/SO/AS-56/2011], revealed about 87 new species of parasitic monogenoids from 45 species of fish representing 31 genera, 13 families, and six orders. This confirms Arunachal Pradesh’s status as a hotspot for fish parasites. Over the last couple of years, a few Arunachali students have developed an interest in fish parasites, partly due to the motivation provided by one of us (AT) and partly due to the growing recognition of the relevance of parasitology in the fisheries and aquaculture sector, which represents the most dominant form of self-employment in Arunachal Pradesh. If these students can secure a permanent position at a state college or university, they can begin a new tradition in systematic work on parasites in Arunachal Pradesh.
{kind=link}