The expression of RAB34 is upregulated in glioma
RAB34 expression data was abstracted from the TCGA and CGGA data resources. We found that RAB34 expression was remarkably higher in GBM (WHO IV) in contrast with that in WHO grade II, as well as grade III gliomas (Fig. 1a, b). Besides, the increased expression of RAB34 was positively correlated with advanced WHO grade of glioma. To further confirm these results, we examined RAB34 expression level in glioma samples, as well as cell lines via qRT-PCR. The data illustrated that RAB34 expression was dramatically increased in glioma samples than that in normal brain tissues (Fig. 1c). Furthermore, GBM samples showed the highest RAB34 expression, which is consistent with above results (Fig. 1d). Moreover, RAB34 expression was upregulated in U251, U87, A172, H4 and SHG44 glioma cells than that in HA1800 (Fig. 1e). These results suggested that the abnormal expression of RAB34 might be involved in glioma progression.
RAB34 expression is enriched in IDH wild-type glioma
Next, we examined RAB34 distribution in four TCGA-defined molecular subtypes. The results revealed that RAB34 was apparently upregulated in classical and mesenchymal subtypes than the other two subtypes in TCGA and CGGA datasets (Fig. 2a, b). Furthermore, we explored the relationship of RAB34 expression with IDH mutation status. We established that IDH wild-type glioma exhibited a higher content of RAB34 in contrast with IDH mutant glioma in TCGA and CGGA datasets (Fig. 2c, d). Subsequently, receiver operating characteristic (ROC) analysis was employed to explore the diagnostic significance of RAB34 as a signature for diagnosing IDH status in glioma. Results showed that areas under curve (AUCs) were 95.1% and 97.6% in TCGA and CGGA cohorts, respectively (Fig. 2e, f). These results suggested that RAB34 was selectively distributed and could serve as a signature for IDH wild-type glioma.
RAB34 expression is negatively correlated with promoter DNA methylation
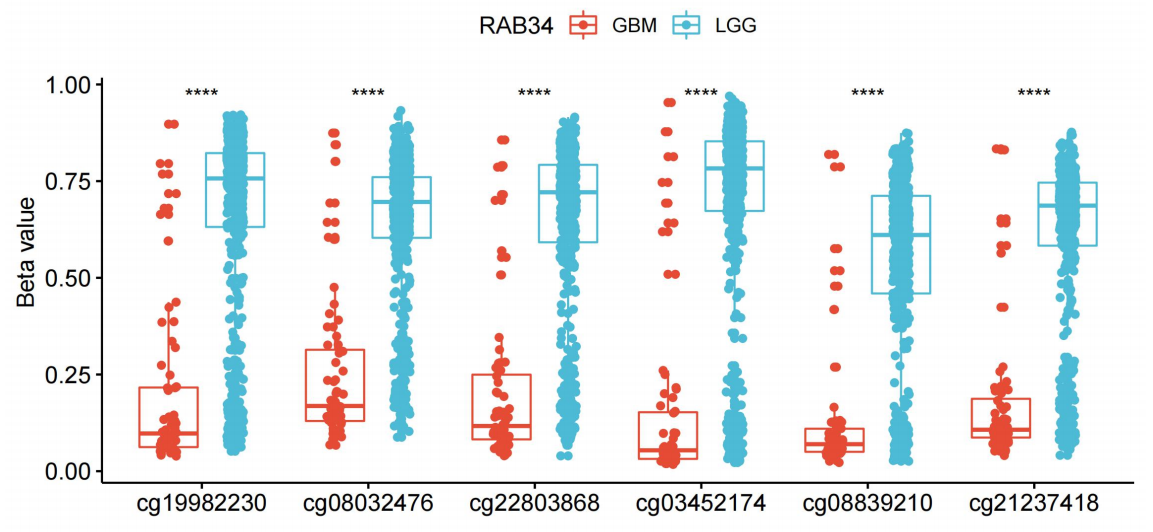
To further investigate the possible mechanism of high RAB34 levels in glioma, we integrated TCGA glioma data sets and identified 594 patients with RAB34 expression along with DNA methylation data. In particular, six CpG sites (cg19982230, cg08032476, cg22803868, cg03452174, cg08839210, and cg21237418) were located in the promoter region of RAB34. Interestingly, we found that the DNA methylation beta values at all these six sites were markedly higher in LGG than that in GBM (Supplementary Figure S1). Additionally, the promoter DNA methylation beta values were increased in the low RAB34 expression group compared with that in the high RAB34 expression group (Fig. 3a). Moreover, all the beta values at promoter region were negatively related to RAB34 mRNA expression (Fig. 3b). To further validate these results, we treated U251 and U87 cells with DAC (decitabine), which is a DNA methyltransferase inhibitor. Western blot results showed that RAB34 expression was increased in response to DAC in a dose of 5µM or 10µM compared with untreated cells (Fig. 3c). Meanwhile, we found that patients who had low methylation levels exhibited a dramatically worse OS in contrast with those who had high methylation levels (Fig. 3d). In summary, these data implied that RAB34 expression was possibly modulated by promoter DNA methylation, and this epigenetic modification could also serve as a promising prognostic signature for patients with glioma.
RAB34 expression is related to immune functions and inflammatory activities in glioma
To identify the biological effect of RAB34 in glioma, GO analysis was conducted. First, RAB34-linked genes in the cohorts were explored with Pearson correlation, with a cut-point of |R| >0.5 along with P < 0.01. Overall, 2220 and 743 positive genes, as well as 1629 and 289 negative genes were detected as remarkably linked to expression of RAB34 in TCGA and CGGA cohorts, respectively. Then, we investigated the biological roles of these genes with the DAVID data resource. The results demonstrated that the genes positively correlated with RAB34 in TCGA dataset were enriched in extracellular matrix organization, immune response, angiogenesis, and inflammatory response (Fig. 4a). Meanwhile, the positive genes in CGGA dataset were enriched in positive modulation of I-kappaB kinase/NF-kappaB signaling, cell-cell adhesion, extracellular matrix organization, and response to hypoxia (Fig. 4b).
To further examine if RAB34 is associated with inflammatory response, seven clusters (including 104 genes) that were defined as metagenes were selected to represent diverse kinds of inflammation and immune response. RAB34 expression was positively linked to most of the clusters except for IgG in TCGA and CGGA datasets (Fig. 4c, d). To verify these data, Gene Sets Variation Analysis (GSVA) was employed to convert these metagenes’ expression levels into enrichment scores, and then corrgrams were generated on the basis of the Pearson correlation values between RAB34 and seven metagenes. As illustrated in Fig. 4e and f, RAB34 expression was positively related to HCK, LCK, MHC-I, MCH-II, STAT1, and interferon but negatively related to IgG, a marker for B cells. Taken together, these results indicated that RAB34 played an important role in the inflammatory response in glioma.
RAB34 expression is correlated with stromal cells and immune cells in glioma
Glioma tissues include not only glioma cells, but also non-tumor cells associated with gliomas, e.g., stromal cells along with immune cells. These non-tumor cells dilute the purity of glioma cells and serve a vital role in glioma biology[26]. Previous researches reported that tumor purity was closely related to major clinical, as well as molecular characteristics, and low-purity cases were more likely to be diagnosed as malignant tumors and independently associated with shorter survival time[27]. Considering the GO analysis data, we next investigated whether RAB34 expression was linked to the infiltrated cells in glioma using ESTIMATE algorithm method described by Yoshihara[28]. The results demonstrated that RAB34 was remarkably related to the stromal score, immune score, and ESTIMATE score in both TCGA and CGGA datasets (Fig. 5a, b). We further employed the CIBERSORT algorithm[29] to investigate the relative proportions of 22 subpopulations of infiltrating immune cells in glioma tissues from TCGA cohort. A comparative summary of the percentages of immune cells in different RAB34 expression groups was briefly depicted in Fig. 5c. Notably, we identified that the percentages of M0 and M2 macrophages, and T cells CD4 memory resting were dramatically elevated in the high RAB34 expression group, while monocytes and plasma cells were remarkably enriched in the low RAB34 expression group. Next, we analyzed the relationship of RAB34 expression with 22 non-malignant cell types. The results showed that 10 types were positively related to RAB34 expression, whereas 6 types were negatively related (Fig. 5d). These findings strongly indicated that RAB34 played a crucial role in the infiltration of stromal cells and immune cells in the glioma microenvironment.
RAB34 expression is related to immune checkpoints in glioma
Immune checkpoint inhibitors designed to restore tumor-induced immunosuppression have become effective anticancer therapies[30]. Therefore, we explored the relationship of RAB34 with immune checkpoints that have been investigated in clinical trials or clinical situations, including PD-1, PD-L1, CTLA4, PD-L2, TIM-3, and B7-H3. Intriguingly, RAB34 exhibited closely positive relationship with these immune checkpoints in all grade gliomas in TCGA and CGGA cohorts (Fig. 6a, b). Similar findings were observed in GBM samples (Fig. 6c, d). These results suggested the possible modulatory influences of RAB34 on these checkpoint members.
Suppression of RAB34 inhibits the biological function and EMT of glioma cells
In order to verify the functional roles of RAB34 in glioma, small interfering RNA si-RAB34 and negative control (si-NC) were inserted into U251 and U87 cells via transfection, respectively. CCK-8 assays showed that the viability of glioma cells was remarkably reduced after the expression of RAB34 was inhibited (Fig. 7a, b). EdU assays also illustrated that the number of cells excited by red light in the si-RAB34 group were much less than that in the si-NC group (Fig. 7c, d). In the transwell assays, we found that the reduction of RAB34 dramatically repressed the migration and invasion abilities of U251 and U87 cells (Fig. 7e).
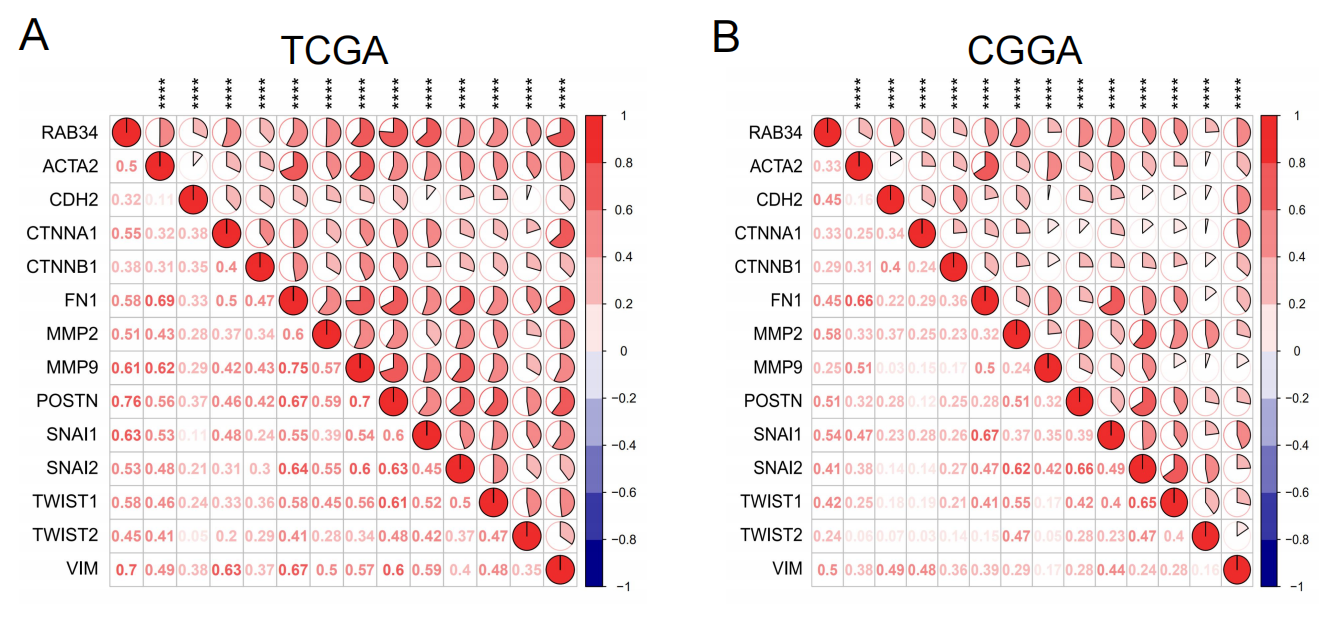
Next, we evaluated the relationship between RAB34 and EMT. Consistently, we found that RAB34 expression was dramatically linked to common EMT biomarkers based on TCGA and CGGA datasets (Supplementary Figure S2A and B). Furthermore, we conducted Western blot assays to assess the influence of RAB34 on the EMT process in glioma cells. As shown in Fig. 7f, si-RAB34 reduced the expression of N-cadherin along with that of Vimentin, but elevated the expression of Claudin-1 in U251 and U87 cell lines. These results revealed that RAB34 may contribute to the EMT progression of glioma.
Overexpression of RAB34 indicates worse prognosis for glioma patients
Considering that RAB34 expression is abnormally expressed in glioma and correlates with histological grade, as well as molecular subtype, we speculated that RAB34 may be an indispensable prognostic signature for glioma patients. Thus, Kaplan-Meier curves were generated to explore its prognostic value. As illustrated in Fig. 8a and b, patients with higher RAB34 expression had a dramatically shorter OS in contrast with patients with lower RAB34 expression in TCGA and CGGA cohorts. Simultaneously, we performed univariate and multivariate analyses to explore whether RAB34 could be an independent predictive factor for glioma patients. Univariate analysis indicated that RAB34 expression was remarkably correlated to OS (Fig. 8c, d). In multivariate analysis, RAB34 expression was still a remarkable prognostic factor after correcting for other clinical factors, such as age, gender and glioma grade (Fig. 8e, f). In summary, RAB34 was an independent predictive factor for patients with glioma.
{kind=link}
{kind=link}