Aphids can damage plants through sugar withdrawal by hijacking sugar metabolism and transport genes to increase their sugar sucking ability (Sun et al., 2019). Plants block the access of aphids to sugars as an active self-protection mechanism (Åhman et al., 2019). This strategy can potentially be exploited to control aphids via the CRISPR/Cas9 editing of genes involved in phloem sugar transport.
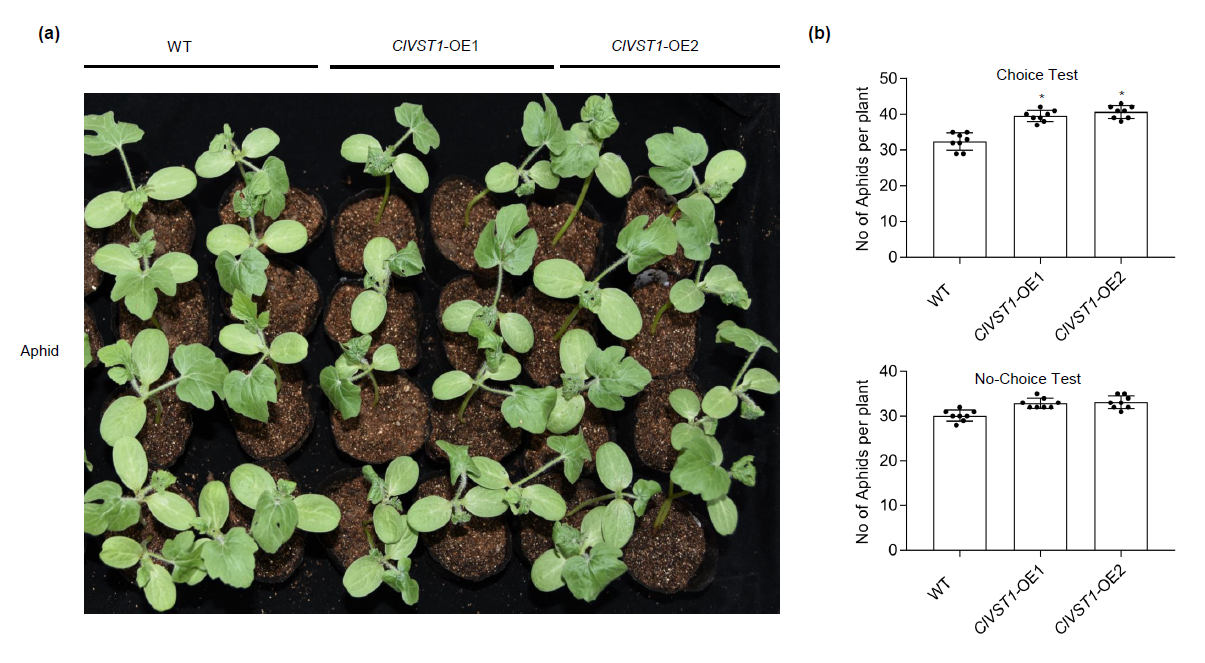
Recently, we reported that a shift in the localization of a vacuolar sugar transporter (VST1) from the tonoplast to the plasma membrane in phloem can contribute to phloem sucrose and glucose unloading in sweet watermelons (Citrullus lanatus) (Ren et al., 2020). Herein, we tested whether the mutation of VST1 can increase tolerance to aphids after aphid feeding. Surprisingly, the emerging leaves of wild-type (WT) plants, but not those of vst1 mutants (vst1-1 harboring a 118-nt deletion and vst1-2 harboring a C/T deletion generated with CRISPR/Cas9 technology) (Ren et al., 2020), showed leaf curling and a large amount of sticky honeydew following aphid infestation in the greenhouse under natural conditions (Figure 1a). To confirm the involvement of VST1 in aphid tolerance, we examined aphid feeding performance on ClVST1 overexpressing (OE) watermelon plants (Ren et al., 2020). Remarkably, the OE plants showed severe leaf curl phenotype than WT plants after aphid infestation (Figure S1a), suggesting that the sugar transporter VST1 plays a negative role in aphid tolerance. Plant defense response to aphid infestation is often reflected by reduced offspring production (antibiosis) in a no-choice test with reduced feeding or by nonpreference (antixenosis) in a choice test (Lei et al., 2014). We performed these tests on different CRISPR and transgenic lines to assess their antibiotic and antixenotic resistance. Approximately twice as many aphids preferred WT versus vst1 plants (Figure 1b); and ClVST1 OE-1 and OE-2 plants showed the most susceptible to aphids in both choice and no-choice tests (Figure S1b). Honeydew production serves as an indicator of insect feeding activity, and honeydew stained by ninhydrin appears as purple spots (Lei et al., 2014). Consistent with the statistical results for the aphid population, the vst1 mutant displayed lighter discoloration of filter paper following ninhydrin staining (Figure 1c), indicating that less honeydew secretion was induced by aphids on vst1 plants than WT plants. This finding was later confirmed by data from quantification based on reading the optical density at 500 nm (Figure 1d). Insect infestation is often accompanied by the accumulation of reactive oxygen species (ROS) in host plants (Lei et al., 2014). We found that upon aphid infestation, vst1 mutant leaves showed less H2O2 accumulation according to 3,3’-diaminobenzidine (DAB) staining (Figure 1e) and less cell death according to the trypan blue staining assay (Figure 1f). Collectively, these data indicate that vst1 mutation can mitigate the damage caused by aphids in watermelon.
We further investigated the transcript levels of VST1 upon aphid infestation by qRT-PCR. The expression level of VST1 was increased at 12 hours post-infestation (hpi) by aphids (Figure 1g). Which further confirmed that VST1 plays an important role in aphid control. Sucrose serves as the optimal carbohydrate substrate for aphid growth and propagation (Hewer et al., 2010). To determine whether VST1-mediated sugar transport is necessary for aphid infestation, we detected the contents of sugars in phloem sap from WT and vst1 mutants by using gas chromatography-mass spectrometry (GC-MS). The levels of both sucrose and glucose were higher in WT plants than in the vst1 mutant (Figure 1h), indicating that sucrose and glucose are important carbohydrate substrates in phloem sap that are transported by VST1 for aphid propagation and infestation. Altogether, our results demonstrated that the mutation of VST1 by genome editing can decrease aphid performance on watermelon by hindering the sugar supply obtained by aphids from phloem sap.
Watermelon is among the most commonly produced fruits around the world, and its production is limited by various insects, especially aphids, which survive on sugars in the phloem sap of their host plants (Clarke et al., 2020). In this context, considered together with the findings of a previous disease study, blocking insect access to nutrients has emerged as an exciting strategy for improving disease and insect resistance in plants (Oliva and Quibod, 2017). Here, we presented the first evidence demonstrating that sugar transporter mutants restrict aphid sucking on watermelon and revealed the possibility of improving crop resistance to aphids or other insects by gene editing. Sugar is also necessary for plant growth and development. It is worthwhile to consider how the constant tradeoff between blocking sugar starvation and defense can be modulated to avoid compromising plant yields under a long-term breeding strategy (Zaidi et al., 2018). Additionally, the fruit sink has the independent capacity for accumulating sugars without being influenced by leaf, stem, root or rootstock tissues in watermelon (Ren et al., 2021). A potential solution to minimize the negative effect of VST1 on sugar content and yield is to perform tissue-specific editing via the precise CRISPR-mediated strategy in the future (Zhu et al., 2020).
{kind=link}