PBOO initiates a pathophysiological cascade of events in the bladder wall, including inflammation, hypoxia, collagen deposition, smooth muscle hypertrophy, apoptosis, and fibrosis. The obstructed bladder undergoes modification with respect to its structure to compensate for the increased resistance to flow, and significant hypoxia ensues owing to the development of high resistance to flow and consequent high intravesical pressure (1–4). Studies in 2 week-obstructed rabbit models revealed the existence of a larger capacity and lower intravesical pressure and compliance of the bladder (2,4). Additionally, studies in Sprague-Dawley rats have shown an increase in end-filling pressure, residual volume, and bladder capacity, as well as a decrease in maximal voiding pressure and bladder compliance (2,10). Some parameters such as voided volume vary with the duration of obstruction time. Yuan et al. revealed that voided volume increased in 3 week-obstructed rats and decreased in 6 week-obstructed group (3). Bladder strip contractility tension and tension sensitivity reportedly decrease in various animal models, including mice, rats, and rabbits (4,10,11).
Presently, there exists a lack of availability of effective treatments for the improvement of bladder function after long-term moderate to severe pBOO in clinical practice. Studies have confirmed the effect of MSCs on the bladder function of pBOO animal models based on the potential applicability of stem cells in tissue reconstruction and organ repair engineering (5, 12–14). However, the mechanism has not been explored at the genetic and molecular levels. Additionally, the disadvantages of MSCs, including the invasive extraction process and high cost, have limited their future clinical applications. Compared with MSCs, USCs are deemed advantageous owing to the simple, noninvasive, and low-cost harvesting methods. Studies have revealed the nephron-protective effect of USCs on renal function realized via anti-inflammatory, antioxidative stress, and antifibrotic activities in acute and chronic kidney disease (15,16). Moreover, USC-exosomes have exhibited pro-neurogenesis effects in the rat brain after the occurrence of ischemic stroke and have demonstrated the exertion of pro-angiogenesis effects and improvement in erectile dysfunction in diabetic rats, thereby exhibiting bladder protection functions (17–19). Presently, there are no reports available on the effects of USCs on the bladder function in patients with pBOO.
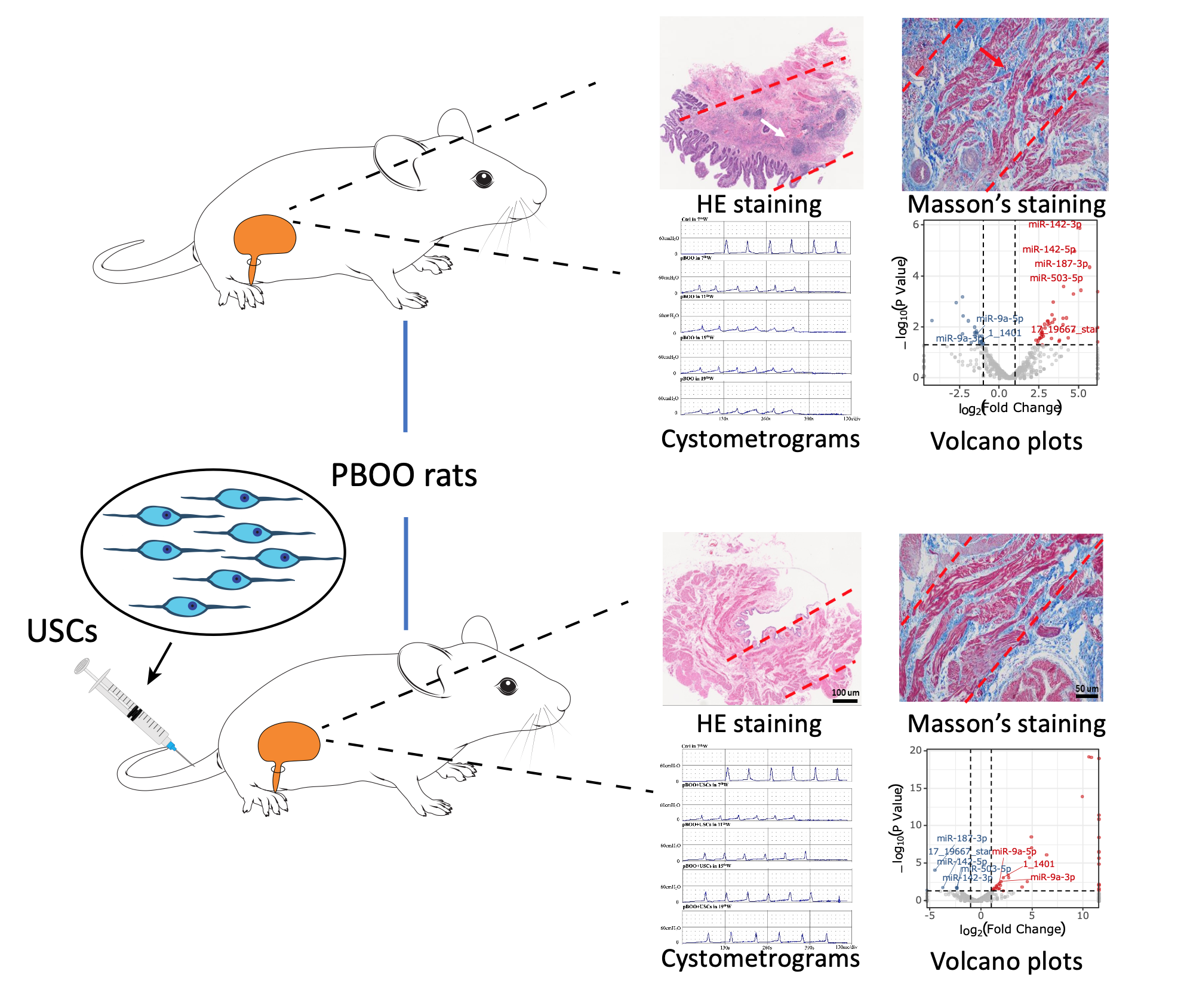
With the aim to explore the effect of USCs on bladder function and morphology in moderate or severe pBOO, we designed a long-term pBOO rat model and dynamically observed the changes from 7- to 19-week-obstructed rats. Similar to the findings reported in previous studies, we observed elevated end-filling pressure, residual volume, and bladder capacity and declined maximal voiding pressure, voided volume, and bladder compliance in the pBOO group compared with the control group. Additionally, once a low-compliance bladder was established, cystometric parameters did not improve without intervention, even if the urethral ligature was removed. Certain parameters, including end-filling pressure, residual volume, and bladder capacity, consistently increased after ligature removal, which consequently resulted in worsening of bladder function. Similar to the case observed in most patients with moderate to severe long-term prostatic hyperplasia, the lower urinary tract symptoms of patients with pBOO cannot be relieved even if there is acceptance of resection of the prostate to relieve bladder outlet obstruction. After subjection to treatment with USCs, few cystometric parameters were significantly improved, including elevated bladder compliance and maximal voiding pressure, declined end-filling pressure, and voided volume. Although residual volume and bladder capacity did not show recovery, there was no further deterioration after subjection to USC treatment compared with the pBOO group. The detrusor muscle contractility and carbachol sensitivity were also improved after USC treatment. Results from the histological analysis and TUNEL assay revealed that the improvement of these functions was mainly related to the reduction of collagen deposition in the extracellular matrix and lowered muscle cell apoptosis.
Furthermore, the urinary bladder was exposed to mechanical stress and was subjected to treatment with USCs, which undoubtedly resulted in modification of gene and protein expression profiles in epithelial and smooth muscle cells, consequently altering the ultrastructure and physiology of cells in the bladder wall. To elucidate the molecular mechanisms underlying this complex process, we conducted bioinformatics analysis of miRNA–mRNA expression profiles of rats in the three groups. We focused on the numerous miRNAs and mRNAs that were significantly and differentially expressed in the bladder following pBOO; those that underwent reversed expression following USC treatment were examined. We identified miR-142 and miR-9a as the two largest nodes of DEMs in the miRNA–gene interaction network.
MiR-142-3p and miR-142-5p are reportedly overexpressed in the bladder tissue of pBOO mice (20). A previous study confirmed that miR-142-5p is overexpressed in the bladder of patients with pBOO (21). MiR-142-3p and miR-142-5p influence inflammation and immune response, the levels of which have been shown to decrease in mouse models of cardiac hypertrophy. miR-142-5p targets acetyltransferase p300 and is implicated in cardiac growth; additionally, an increase in miR-142 expression during cardiac growth is critical for cell survival (22,23). The in vivo inhibition of miR-142-5p with locked nucleic acid-modified oligonucleotides reduces CCL4-induced liver fibrosis and bleomycin-induced lung fibrosis in mice. Furthermore, macrophages derived from the tissue samples of patients with liver cirrhosis and idiopathic pulmonary fibrosis display increased levels of miR-142-5p (24). Therefore, it can be inferred that miR-142-5p regulates macrophage profibrogenic gene expression during chronic inflammation. In this study, miR-142-3p and miR-142-5p were overexpressed in pBOO rats, whereas their expression was suppressed by USCs, indicating that miR-142 participated in the antifibrosis effect of USCs in pBOO rats.
Studies have revealed that miR-9a participates in anti-inflammatory and anti-apoptotic functions and is related to neurogenesis. The downregulation of miR-9a-5p expression may increase the levels of NLRP1 inflammasome proteins, cleaved caspase-1, interleukin (IL)-1 b, and IL-18. In contrast, the overexpression of miR-9a-5p markedly prevents the abnormal expression of these proteins and is accompanied by the downregulation of NLRP1 expression in ischemia-like conditions, both in vivo and in vitro (25). Further, miR-9 prevents cardiac dysfunction and inhibits cardiomyocyte apoptosis in myocardial infarction mouse hearts (26). However, a study indicated that miR-9a-5p is a novel regulator of HSCs in the setting of increased pressure and hepatic fibrosis in rats and that the overexpression of miR-9a-5p following hepatic stellate cell activation perpetuates the fibrotic response (27). In our study, we found that both miR-9a-3p and miR-9a-5p levels were decreased in pBOO rats and increased in USC-treated pBOO rats. They may confer protection to bladder functions by reducing the inflammatory response of bladder tissue and reducing detrusor cell apoptosis, aspects which should be confirmed through further studies.
To obtain insights into the classification of miRNAs, we conducted GO and KEGG analyses to predict the target genes and possible pathways. The results revealed various GO terms associated with superoxide-generating NADPH oxidase activity, NADPH oxidase complex, semaphorin receptor activity, semaphorin–plexin signaling pathway involved in axon guidance, oxidoreductase activity, acting on NAD(P)H, oxygen as an acceptor, and hydrogen peroxide biosynthetic process. Additionally, it was observed that various pathways were involved, including leukocyte transendothelial migration, chemokine signaling pathway, cytokine–cytokine receptor interaction, necroptosis, NOD-like receptor signaling pathway, and apoptosis. The necroptosis and cytokine–cytokine receptor interaction signaling pathways may be responsible for the pathological alterations observed in this study and may help provide insights for the formulation of a strategy for therapeutic intervention, following further investigation. The necroptosis signaling pathway is a programmed form of necrosis. It can be initiated by different stimuli, such as tumor necrosis factor (TNF) and TNF-related apoptosis-inducing ligand. It has previously been demonstrated that cytokine and immune response pathways, transforming growth factor, nitric oxide signaling, and hypertrophic PI3K/AKT signaling pathways are shared among all pBOO phenotypes. AP-1 and NF-kB are considered the dominant transcription factors, and TNF-α is recognized as the top upstream regulator (21). However, classic fibrosis-related pathways such as the transforming growth factor beta signaling pathway were not identified in this analysis. Therefore, it may be implied that USCs may inhibit the initiation of the pathophysiological cascade of events in the bladder wall via the necroptosis and cytokine–cytokine receptor interaction signaling pathways. Additional research is warranted to verify the exact roles of the identified miRNAs, genes, and signaling pathways in the pathogenesis of bladder remodeling induced by pBOO.
{kind=link}