Mealworm beetles
Mealworm beetles are stored grain product pests that live several months in populations of variable density and at sex-ratio of about 50% (± 20%). T. molitor males and females may initiate reproduction from the fifth day post emergence, although they reach their full sexual maturity from the eighth day post emergence. They can mate many times with several partners within their 2 to 5 months of adult life. Females are continuously receptive to mating during adulthood and may produce up to 30 eggs per day although egg production may decline after three weeks[43]. Although able to store sperm in their spermatheca, females need to mate frequently to maintain high egg production [44].
The immune system of T. molitor relies on both constitutive cellular (e.g. haemocytes) and enzymatic (e.g. prophenoloxidase system) components at the core of the inflammatory response [45]. Their activity is cytotoxic [46], causing self-damage [47] and lifespan reduction [48–51]. They were found to decrease after mating [52] and either decline [53] or increase [54] with age. In addition, the inducible production of antibacterial peptides in the haemolymph [45] is an energetically costly process that may reduce survival [55]. As selection on immune expression and immune regulation might be weaker after reproductive senescence, age-related decline of baseline levels of immunity might be observed and immune activation may occur at old age due to dysregulation [54, 56].
Artificial populations and experimental design
Virgin adult beetles of controlled age (10 ± 2 days post-emergence) were obtained from pupae haphazardly sampled from a stock culture maintained in laboratory conditions (24 ± 2°C, 70% RH in permanent darkness) at Dijon, France. Prior to the experiments, all these experimental insects were maintained separately in laboratory conditions, and supplied ad libitum with bran flour and water, supplemented by apple.
Fifteen artificial populations of 100 adult beetles were made according to three sex-ratio conditions. Five populations had a balanced sex-ratio, each comprising 50 males and 50 females (thereafter named the 50%_males condition), and were considered as the reference populations. Five populations had a male-biased sex-ratio, each comprising 75 males and 25 females (75%_males). Finally, five populations had a female-biased sex-ratio, each comprising 25 males and 75 females (25%_males). Each population was maintained in a plastic tank (L x 1 x H, 27 x 16.5 x 11.5 cm) containing bran flour, supplied once a week with apple and water. Every two weeks, each population was transferred into a clean tank supplied with fresh bran flour, thus avoiding the development of the progeny with the experimental adults.
Age specific reproductive assay
Reproductive capacity of females and males in each population was estimated weekly. To this purpose, 4 females haphazardly picked in each population were individually transferred into a plastic Petri dish (9 cm in diameter), containing bleached flour, a 2 mL centrifuge tube of water and a piece of apple. Each female was allowed to lay eggs in the Petri dish for 3 days, and was then returned to their initial population box. Two weeks later, the number of larvae was counted in each Petri dish to quantify female fertility, which is the number of viable larvae produced per female [57].
Concomitantly, four males haphazardly picked in each population were also individually transferred into Petri dishes, as above. Reproductive success of males was estimated through direct measures of their fertility (number of viable offspring per male [57]) instead of measuring spermatophores or counting the sperm, which are rough surrogates of male reproductive success. Each male was provided with a virgin female aged from 8 to 15 days for 24 hours and was then returned in its initial population. Each female was then allowed to lay eggs in the Petri dish for three additional days to estimate, as described above, male fertility. In T. molitor, males may affect female fecundity (number of potential eggs produced by the female) and therefore their fertility, according to the respective quality of spermatophores and sperm transferred during mating. Consequently, male’s success was a measure of the potential reproductive effort, not the one realized within its experimental population.
While assayed for their reproduction, focal females and males were replaced by marked individuals of the same age and sex in all the populations, to keep sex-ratio and density constant. Substitutes were from the same cohort as the experimental insects, kept in a separate tank of mixed-sex population. They were marked by clipping a piece of one elytra. When focal insects assayed for their reproduction were returned into their initial population box, substitutes were removed and returned into their tank.
Estimation of male and female reproductive effort at the population level
Survival of the insects was checked weekly, and dead insects were replaced by marked substitutes of the same sex and about the same age to keep the population sex-ratio and density of individuals constant. No measurement was performed on these marked individuals.
As the experimental design does not allow gathering measurements of longevity and fertility for each individual of the population, we estimated male and female reproductive effort (RE) at the population level, from the above age-specific measures of fertility, for each of the five population replicates, within sex-ratio conditions. This estimate was calculated as the total number of viable larvae produced per female or male in each replicate (i.e. the cumulative number of larvae produced during the whole experiment in a given replicate divided by the number of females or males tested for this replicate), divided by their respective average lifespan in the population replicate (i.e. the average lifetime of females or males in each relicate). The equation is given as follow:
See equation 1 in the supplemental files.
Where l is the total number of offspring (here viable larvae) produced per assayed females or males in the population replicate r, and ML is the recorded mean lifespan (in weeks) of males and females in the replicate r. RE values (as offspring per individuals and per mean weeks of survival in the population) of each sex and population replicate within each sex-ratio condition were used as data points for comparisons between modalities of sex-ratio.
Note that female RE values are likely representative of both female and male conditions resulting from the experiment, because the female reproductive performance resulted from mating with males from their respective population. By contrast, male RE values are representative of the male condition only, because male reproductive performance was standardized by pairing it with a virgin and age-controlled female that did not experience the experimental conditions. Therefore, male RE must be seen as a surrogate of male reproduction potential.
Body condition and haemolymph collection
At weeks 2, 4, 6 and 12 after the start of the experiment, 4 females and 4 males were picked at random in each population to estimate their body condition and immunity. The first three time points were chosen as being relevant of the time period during which most of the beetle reproduction is achieved and survival is still relatively high [44]. It is also within this period of time that a potential decline in somatic protection, including immunity, is predicted. The last time point corresponds to a period of time when reproduction should have almost ceased and when few beetles remain alive. As immunity measurements was destructive sampling, sampled insects were replaced by marked substitutes as above, to keep sex-ratio and density constant. However, this substitution was definitive, as sampled individuals were not returned to their initial population box after being assayed. The below estimation of the insect body condition and immunity was done as described in [58]. Beetles were first sized by measuring the length of the right elytra with Mitutoyo digital callipers (precision 0.1 mm) and weighed to the nearest mg with an OHAUS balance (discovery series, DU114C). Body condition was then estimated by the residuals of the regression between body size and body mass. Then, beetles were chilled on ice for 10 min before the sampling of 5 µL of haemolymph from a wound made in the beetle’s neck and flushed in a microcentrifuge tube containing 25 µL of phosphate-buffered saline (PBS 10mM, pH 7.4). A 10-µL aliquot was immediately used to measure haemocyte count. Another 5-µL aliquot was kept in an N-phenylthiourea-coated microcentrifuge tube (P7629, Sigma-Aldrich, St Louis, MO, USA) and stored at -80 °C for later examination of its antibacterial activity. The remaining haemolymph solution (15 µL) was further diluted in 15 µL of PBS and stored at -80 °C for later measurement of its phenoloxidase activity.
Immune parameters
Haemocyte count was measured using a Neubauer improved haemocytometer under a phase-contrast microscope (magnification x 400).
Antimicrobial activity of the haemolymph was measured using the inhibition zone assay described in [58]. Briefly, an overnight culture of the bacterium Arthrobacter globiformis from the Pasteur Institute (CIP105365) was added to a Broth medium containing 1 % agar to achieve a final concentration of 105 cells.mL-1. Six millilitres of the medium was subsequently poured per Petri dish and, after solidification, 12 wells were made inside the agar plate in which 2 µL of each haemolymph sample was deposited. Plates were then incubated overnight at 28 °C and the diameter of each zone of inhibition was measured.
For each haemolymph sample, both (i) the activity of naturally activated phenoloxidase (PO) enzyme only (hereafter PO activity) and (ii) the activity of PO plus that of proenzymes (proPO) (hereafter Total-PO activity) were measured using the spectrophotometric assay described in [59]. Total-PO activity quantification required the activation of proPO into PO with chymotrypsin, whereas PO activity was measured directly from the sample. Frozen haemolymph samples were thawed on ice and centrifuged (3,500 g, 5 min, 4 °C). In a 96-well plate, 5 µL of supernatant were diluted in 20 µL of PBS and were added either 140 µL of distilled water to measure PO activity or 140 µL of 0.07 mg. mL-1 chymotrypsin solution (Sigma-Aldrich, St Louis, MO, USA, C-7762) to measure Total-PO activity. Subsequently, 20 µL of a 4 mg.mL-1 L-Dopa solution (Sigma-Aldrich, St Louis, MO, USA, D-9628) were added to each well. The reaction proceeded for 40 min at 30 °C, in a microplate reader (Versamax, Molecular Devices, Sunnyval, CA, USA). Reads were taken every 15 s at 490 nm and analysed using the software SOFT-Max®Pro 4.0 (Molecular Devices, Sunnyvale, CA, USA). Enzymatic activity was measured as the slope (Vmax value: change in absorbance unit per min) of the reaction curve during the linear phase of the reaction and reported to the activity of 1 µL of pure haemolymph.
Statistics
Cox-regressions with a time-dependent covariate were used to analyse the difference in survival rates with respect to sex-ratio during the time (in weeks) from the start of the experiment and the death of all individuals. Sex-ratio was coded as categorical variables. The effect of sex ratio in the statistical model used the reference survival function generated from the data derived from the females or the males of the 50%-male sex-ratio condition. Time (in weeks) was incremented as a covariate in interaction with sex-ratio in the model as hazard ratios when the survival functions where not constant over time (for more details, see [60]). The analyses of fertility (i.e. the number of larvae produced per female or male at each week) and immune parameters were performed using mixed models, either Linear or Generalized linear depending on the nature of the data (see table legends). Starting models included sex-ratio condition, week (continuous variable for fertility, ordinal variable for immunity), their interaction, body condition and replicates treated as a random factor (REML estimates of variance component). The models presented here are those minimizing the AICc, where DAICc > 2 is usually considered to be good support [61], after comparisons of all models including predictors and their interactions, in a stepwise fashion (see Table S1). The analyses of reproductive effort were made using ANOVA testing the effect of sex-ratio conditions. Analyses were made using IBM® SPSS® Statistics 19, JMP v. 10.0 and R version 3.3.2 (The R Foundation for Statistical Computing, Vienna, Austria, http://www.r-project.org). All the data files are available from the Dryad data base [62]
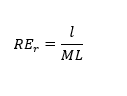{kind=link}