The chlorophyll content is an important parameter for the determination of photosynthesis in plants and is involved in the absorption, transfer and conversion of light energy during photosynthesis. Chlorophyll plays a key role in photosynthesis, and changes in its content reflect the degree of photosynthesis that occurs [23]. The central atom of chlorophyll, magnesium, can be replaced by heavy metals such as cadmium, copper, zinc, lead and mercury, hampering the light-harvesting ability of chlorophyll and impairing photosynthesis performance in stressed plants [24]; furthermore, the chlorophyll in plants under heavy metal stress is subject to metabolic disruption and inhibited synthesis [25, 26]. Zn is an essential element for plant growth, and when the Zn concentration exceeds a critical value, large amounts of Zn enter chlorophyll cells and disrupt chlorophyll metabolism and chlorophyllase activity, leading to a decreased synthesis of chlorophyll. In this paper, the three studied plants showed a decrease in their chlorophyll A and B contents under heavy metal stress from tailings, while the chlorophyll A/B ratio reflected the relative ratio of stacked to unstacked membrane domains and was inversely proportional to the degree of cystoid compression. Cystoid compression is responsible for bringing the light-trapping chlorophyll protein complex (LHCII) closer together, making it useful for maximizing the light-harvesting capacity. Additionally, the PSII reaction center in the stacked region is in the same pigment bed as the adjacent LHCII, which further increases the energy transfer capacity of the membrane [27].The increase in the chlorophyll A/B ratio after heavy metal stress is assumed to be due to the cystoid not being stressed, and the reduction in the cystoid is a positive sign of plant adaptation to environmental stress [28].
Photosynthesis is an important indicator to evaluate plant growth and development and plant productivity. Stomata are important structures in plant leaves that control photosynthesis, CO2 and water uptake and transpiration; thus, the physiological indicators of stomata have an important influence on the photosynthetic physiology of plants. In general, stomatal factors mainly include the number, size and opening of stomata, while nonstomatal factors include enzyme activity and the photosynthetic components. Under environmental stress, both stomatal and nonstomatal limitations lead to a decrease in Pn, but Ci is used to distinguish between stomatal and nonstomatal limitations [29]. When both Pn and Ci are reduced, the decrease in photosynthetic capacity is caused by stomatal limitation; however, when the photosynthetic rate decreases and Ci increases, the cause is nonstomatal limitation [30]. In this paper, Kochia scoparia under heavy metal stress showed a significant decrease in Pn, Gs, and a significant increase in Ci, compared to the control group, indicating that under heavy metal stress, the Kochia scoparia leaf Pn decreased mainly due to nonstomatal limiting factors. In contrast, under heavy metal stress, Chenopodium glaucum and Atriplex centralasiatica showed a significant decrease in Pn, Gs, and Ci compared to the control group, indicating that the photosynthetic rate of these two plants decreased due to stomatal limiting factors. All three plants showed a significant decrease in transpiration rate after heavy metal stress, and the order of decrease was: Atriplex centralasiatica > Chenopodium glaucum > Kochia scoparia.
Studies on the relationship between the stomatal regulation, leaf transpiration rate and heavy metal accumulation in the plants are now well documented [31, 32], and heavy metal accumulation in the aboveground parts of the plants is subject to several processes, including root uptake, radial transport, xylem loading and transport from roots to the aboveground parts. Transpiration is the driving force for a large amount of water flow in this process. Therefore, the heavy metal content of the aboveground part of the plant, is thought to be achieved due to the entire flow transport mechanism from the soil substrate to the stem and then to the leaves through transpiration [33]; this flow of heavy metals is enhanced when the stomatal conductance increases, which increases the flux of water-soluble pollutants to the upper part of the plant. It was hypothesized that plants with more efficient water use and fast stomatal response were more desirable target candidates for tailings remediation. The transpiration rate and hydraulic conductivity of Atriplex centralasiatica were significantly higher than those of the other two plants, and its duct diameter was also larger than those of the other plants. Therefore, Atriplex centralasiatica had the highest heavy metal content among the three plants.
There are two factors influencing the hydraulic conductivity of plants: the intrinsic factors of plants, including the hydraulic structure, morphological structure and osmotic potential of plants; and the extrinsic factors, including soil factors such as the water status, nutrient conditions, salt content, temperature, and atmospheric factors such as the available light, CO2 concentration, and solar radiation. Usually, exogenous factors cause changes in endogenous factors [34]. Plants can regulate the hydraulic properties of xylem under different stress conditions and can maintain a high hydraulic efficiency of stems and roots by reducing the root hydraulic conductivity [35]. Maggio et al. (1995) revealed that heavy metals play a strong inhibitory role on the hydraulic conductivity of tomato roots, and their study showed that the root hydraulic conductivity significantly reduced by 57% during heavy metal ion treatment [36].
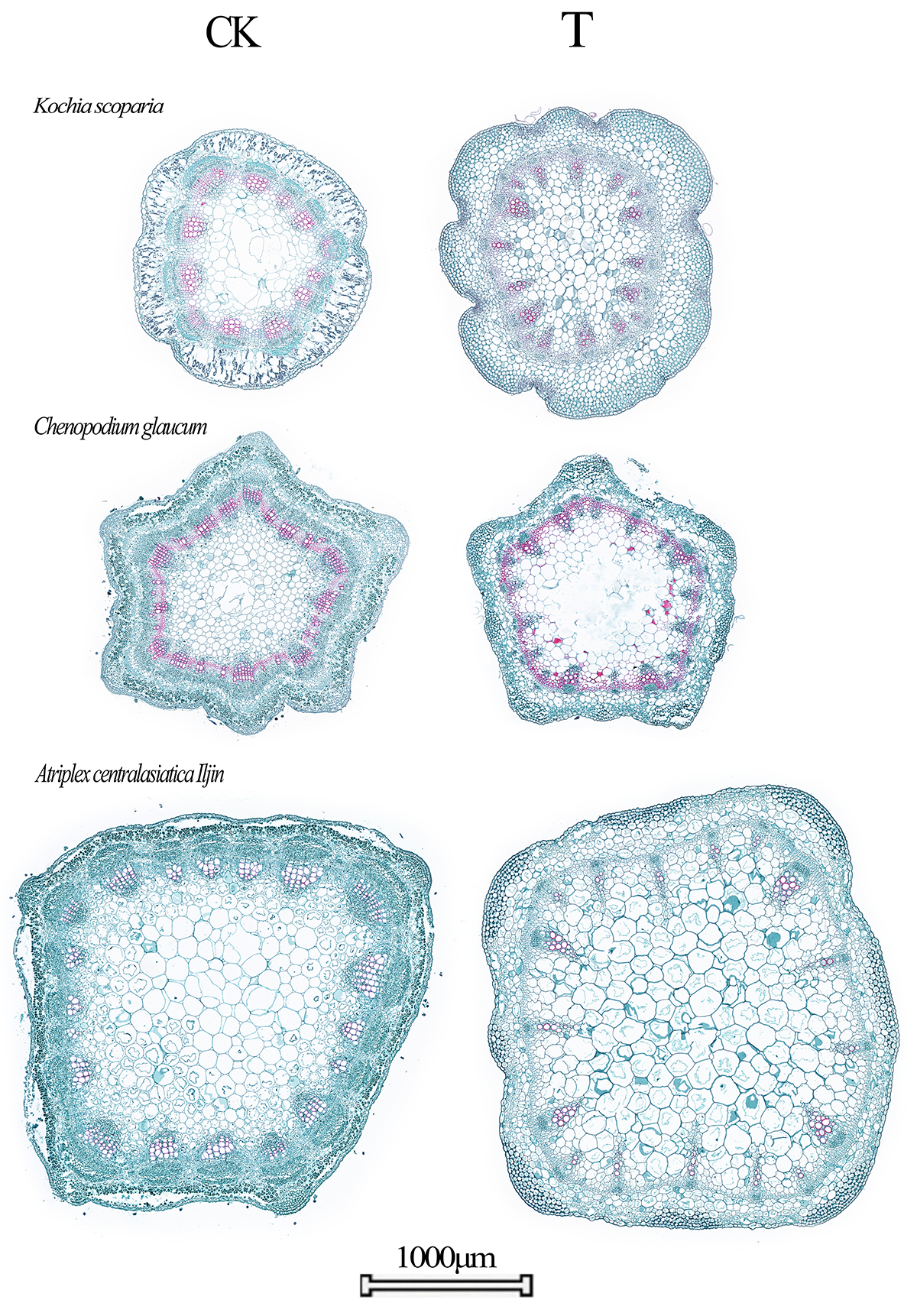
This study discussed the changes in plant stem hydraulics indicators under heavy metal stress, and it was found that all four hydraulics indicators of the three plants showed a significant increase under heavy metal stress, thereby leading to a higher leaf specific conductance. These results are consistent with the results of Adriana M's study on the regulation of the plant response under copper stress, which found an increase in the leaf specific conductance of plants under heavy metal copper stress [37]. The present study showed that the hydraulic conductivity and leaf specific conductance were significantly higher in the three studied plants grown under long-term heavy metal stress than in those grown in the native desert (Fig. 4).
Most of the current studies on plant stem anatomy are on drought and salinity stress, and rarely involve heavy metal stress, but there are studies on the enrichment of heavy metals by non-hyperenriched plants. For example, plants can chelate heavy metals through substances such as polysaccharides in the cell wall [38], and the cell wall thickness of mosses can be increased by inducing organizational changes of the cell wall polysaccharides and cell wall structural morphology [39].
In the present study, we found that the duct density and relative sparing area of all three plants under complex heavy metal stress showed a significant decrease compared to the control, while the xylem duct wall showed a trend of thickening. These results were similar to Krzesowska et al., who previously found that the cell wall pectin chelates with heavy metals entering the cell wall, resulting in an increase in cell wall thickness and hardness [40]. This study showed that the duct wall thickness of desert plants was significantly higher than that of the unstressed group after being subjected to long-term heavy metal stress, and the duct wall thickness after stress was positively correlated with the heavy metal content in the stem. Hajihashemi et al. (2020) found that the ductal density of wheat roots decreased during heavy metal effluent treatment [41], and the same trend was found in the stems of our study, which showed that the ductal density of plants under stress was significantly lower than that of the unstressed group.
This increase in duct diameter is consistent with the findings of Adriana et al. (2016), in which purple willow (Salix purpurea L.) showed an increase in its xylem duct diameter after stress by copper ions and other heavy metals [37]. Accordingly, we hypothesize that transpiration is not the only factor that promotes the increase in plant hydraulic conductivity because the stem hydraulic conductivity from the underground to the aboveground parts of plants can be maintained by increasing the vascular diameter when under stress.
{kind=link}