The presence of segmented appendages with joints consisting of hardened (sclerotized) cuticle connected by flexible membranes – formally known as arthropodization – represents the most recognizable character of most extant and extinct euarthropods [01, 02, 06, 07, 08, 13, 21]. Arthropodized limbs are enormously plastic in their shape and function [22], and thus represent an important evolutionary innovation that contributes towards the substantial diversity and ecological versatility that characterizes this phylum. Despite the significance of arthropodization as a synapomorphy of Euarthropoda, there is a lack of consensus in terms of its precise origin among stem-lineage representatives. The phylogenetically earliest evidence of arthropodization is found among radiodonts, diverse nektobenthic stem-group euarthropods that played an important ecological role in early marine ecosystems during the early Phanerozoic [01, 04, 16, 23]. Radiodonts possess a single pair of multiarticulated and arthropodized raptorial frontal appendages that mainly served a feeding function, either for grasping, crushing, filter feeding or sediment sifting. Although some radiodonts also feature robust appendicular “gnathobase-like structures” associated with the functional head region [23], the rest of the body consists of an unarthrodized trunk with metamerically arranged lateral body flaps for swimming [04]. By contrast, the first appearance of fully arthropodized ventral appendages remains contentious. Suggested evidence for arthropodized legs in Cambrian lobopodians [24] has been regarded as preservation artefacts caused by folding of the flexible or partially decayed cuticle [25]. Since the development of fully arthropodized biramous trunk appendages represents one of the major transitions in early euarthropod evolution [01, 02, 03, 08, 09, 13, 15, 22], resolving this issue carries direct implications for understanding the phylogenetic relationships among early representatives, as well as the emergence of one of the most versatile animal body plans during the Cambrian Explosion.
Recent studies suggest that some of the earliest branching euarthropods bore a broad carapace that covered a weakly sclerotized trunk consisting of ring-like tergites, and largely homonomous pairs of biramous appendages [01, 07, 13, 14, 15, 19]. Among this paraphyletic grade of Cambrian bivalved euarthropods, the isoxyids have been repeatedly compared with radiodonts based on the presence of a pair of raptorial frontal appendages [07, 14, 15, 19, 20]. Despite exceptional soft-tissue preservation in isoxyids including the stalked eyes and paired gut diverticulae, the detailed morphology of their body and biramous appendages remains poorly understood, generally obscured by the dorsal carapace covering the entire body [10, 11, 12, 14, 15, 17, 18, 26]. The Burgess Shale Surusicaris has some of the best-preserved biramous trunk appendages in isoxyids described to date [15], interpreted as weakly sclerotized and simple, annulated limbs with an elongate exopod bearing marginal setae. The trunk appendages of Isoxys volucris from Sirius Passet [11] show crudely preserved paddle-shaped exopods, and endopods without clear signs of segmentation. Although the presence of putative podomere boundaries has been suggested for I. curvirostratus and I. auritus from Chengjiang [12, 17], critical details of the appendicular morphology are missing in all cases to fully assess their evolutionary and ecological significance. Furthermore, whether the trunk of isoxyids was fully arthrodized or not remains completely unknown.
Here, we revise the morphology of Isoxys from the early Cambrian (Stage 3) Chengjiang biota in South China. We employ micro-computed tomography (micro-CT) imaging and 3D computer rendering techniques to investigate the exceptionally preserved pyritized three-dimensional organization of Isoxys biramous appendages, and demonstrate that isoxyids had higher degrees of morphological and functional specialization than previously considered [10, 11, 12, 14, 15, 17, 18, 26].
Morphological description
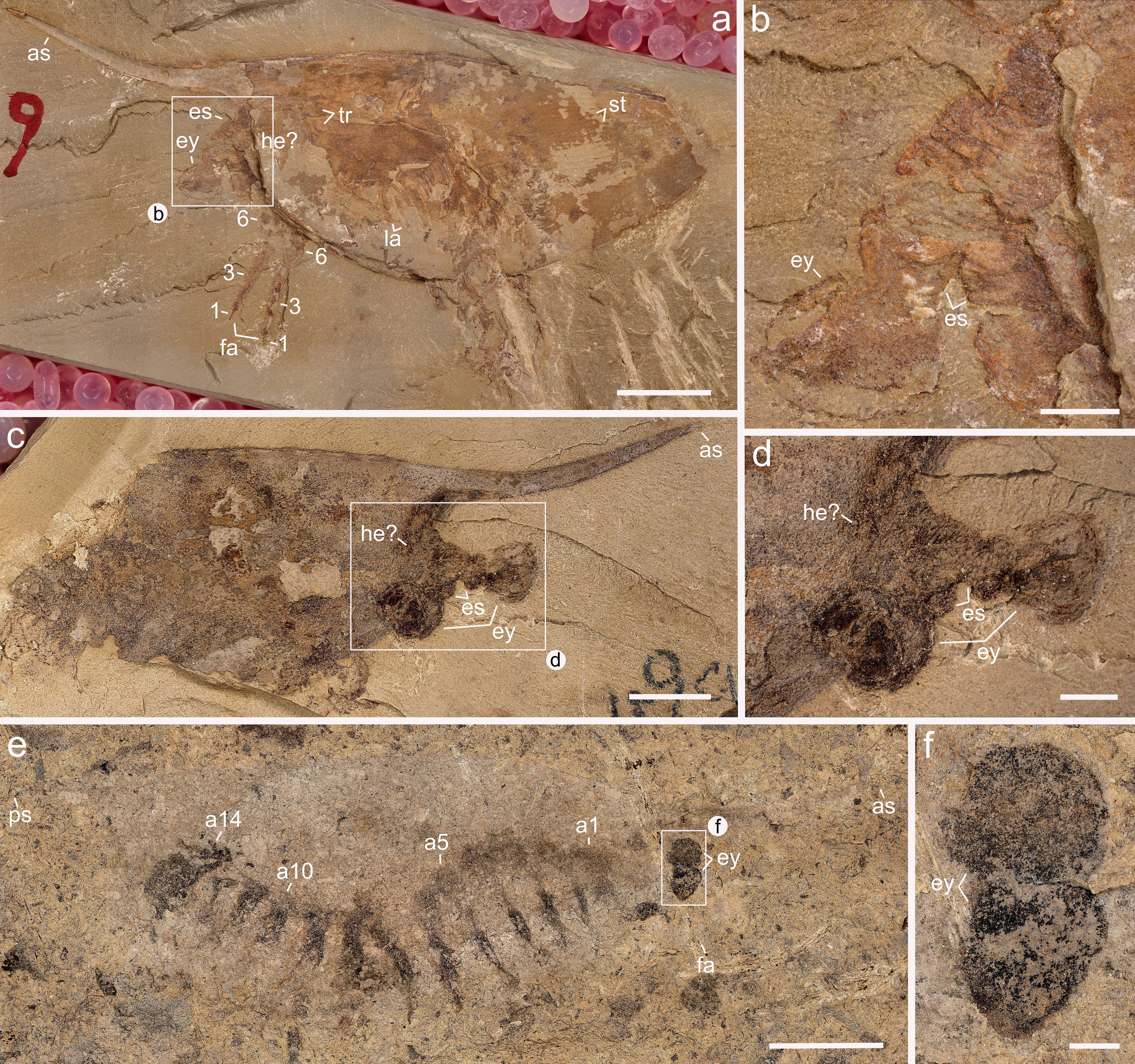
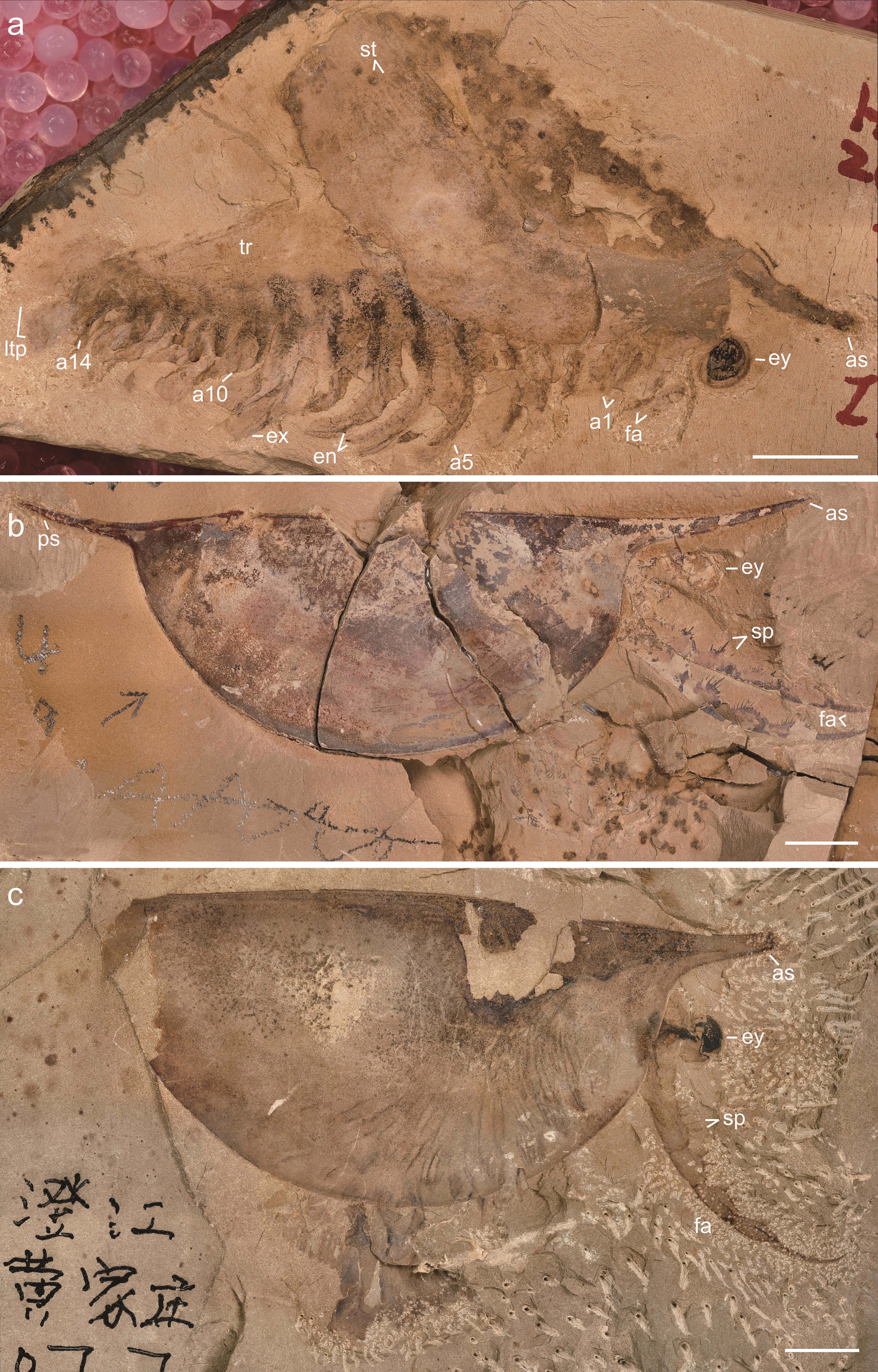
We investigated new material of Isoxys curvirostratus [12] (Fig. 1; Extended Data Fig. 1–4), as well as previously published material of Isoxys sp. [26] (Fig. 2) from Chengjiang. Isoxys species share fundamental aspects of the overall morphology, including the presence of a bivalved dorsal carapace with a semicircular smooth margin, anterior and posterior cardinal spines, prominent stalked eyes and robust frontal appendages [12, 17, 26] (Figs. 1, 2). I. curvirostratus is distinguished by the presence of a convex dorsal carapace margin, upward bending anterior cardinal spine approximately three times longer than the posterior spine, and the presence of longitudinal striations on the posterior part of the carapace (Fig. 1; Extended Data Figs. 1–4). Isoxys sp. cannot be ascribed to an existing species due to the lack of diagnostic characters such as the frontal appendages or carapace ornamentation [12, 17], and thus is treated in open nomenclature following ref. [26]. Carapace length – measured between the bases of anterior and posterior spines – ranges from 24 to 28 mm for I. curvirostratus, and 19 mm for Isoxys sp. in our studied material (Figs. 1, 2). Comparisons with previous reports on Chengjiang Isoxys species [12, 17, 26] suggest that our material corresponds to adults based on their size.
Soft tissues have been described for both I. curvirostratus and Isoxys sp. [12, 17, 26]. The anterior end of the body bears a pair of prominent stalked eyes with a spherical shape, and which protrude beyond the anterior carapace margin directly below the cardinal spine (Figs. 1, 2; Extended Data Figs. 1–4). The appearance of the eyes in Isoxys closely resembles that of the fossilized ocular structures in other Chengjiang euarthropods [2, 4, 6], consisting of a light outer layer and a dark internal mass, which likely correspond to the eye lens and pigmented retina respectively. A pair of well-developed frontal appendages is also found in close association with the stalked eyes on the anterior end of the body (Fig. 1; Extended Data Figs. 3, 4). The frontal appendages appear to attach behind the eyes, similar to Surusicaris [15], but their precise position within the head and relative to the mouth opening remains uncertain in our material due to the coverage by the carapace, as well as in other Isoxys from Chengjiang [12, 17, 26], Sirius Passet [11], Emu Bay Shale [10], and Burgess Shale [14, 18]. Our material of I. curvirostratus preserves the morphology of the frontal appendage in greater detail than previously described specimens [2, 12]. YKLP 16260 and YKLP 16261 shows that the frontal appendage of I. curvirostratus consists of six podomeres that are longer (sag.) than thick (trans.), and follows a distinctive curvature in which the ventral side is facing upwards (Fig. 1m, n). The basal podomere has a subtrapezoidal shape and lacks endites, whereas the following four podomeres are robust, subequal in length (sag.) and have distinctly curved ventral margin that bears up to a dozen spinose endites that are longer towards the podomere midline, and shorter towards the margins (Fig. 1n; Extended Data Fig. 4b). The sixth podomere is a terminal claw, of subequal length to the previous podomeres but with a slender outline. The terminal claw also bears spinose endites that are consistently short and point distally (Fig. 1o). The frontal appendages of I. curvirostratus are morphologically distinct from those of I. auritus, also known from Chengjiang [2, 17], as the latter consists of nine podomeres with a subrectangular outline, with subequal length (sag.), each bearing a single median spinose endite.
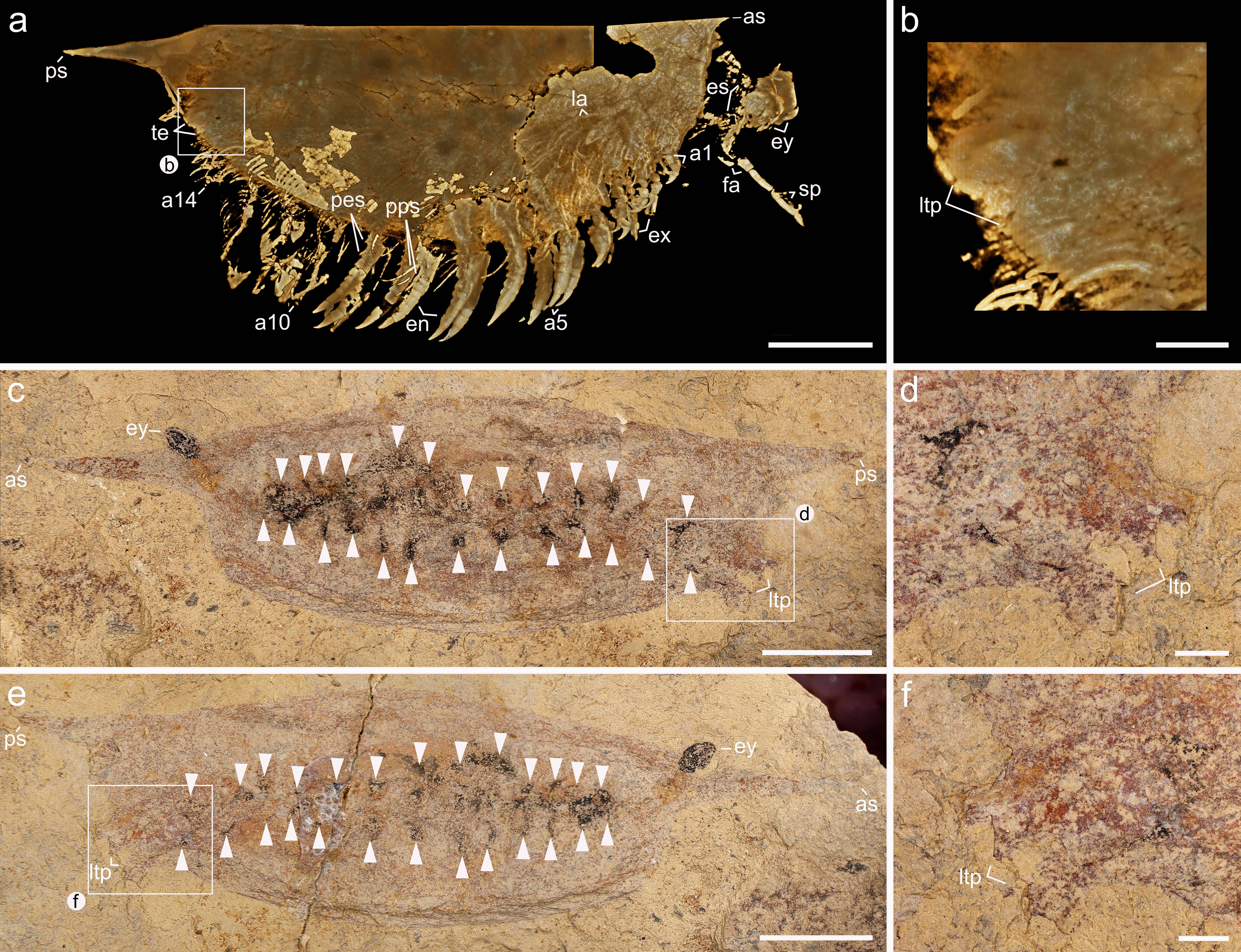
The carapace of Isoxys covers most of the trunk morphology in all specimens described to date [2, 10, 11, 12, 14, 17, 18, 26]. New material of Isoxys curvirostratus with a partially displaced carapace informs the organization of the trunk region (Fig. 1k, l; Extended Data Fig. 4a). YKLP 16261 demonstrates that the trunk of I. curvirostratus clearly lacks any indications of dorsal arthrodization such as well-defined tergites or epidermal segmental boundaries, despite the presence of soft tissues including the eyes, a complete biramous appendage series, and paired telson flaps on the posterior end (Fig. 1l; Extended Data Fig. 4a). The quality of preservation of YKLP 16261 featuring delicate structures such as the stalked eyes and appendages indicates that this lack of arthrodization is legitimate, rather than a taphonomic artifact caused by decay. The overall surface appearance of the biramous appendages in the studied specimens of I. curvirostratus and Isoxys sp. is comparable to those in previous reports [2, 12, 17_Fuetal2014, 26]. The appendages consist of relatively simple endopods and exopods with setae, and appear nearly homonomous, except for a gentle increase in size from the anterior end to the middle of the body, and then decrease in size from the middle to the posterior end (Figs. 1a, l, 2a; Extended Data Fig. 3). Both Isoxys curvirostratus [12] and Isoxys sp. feature 14 pairs of ventral biramous appendages (Figs. 1, 2), which distinguish them from the 11 pairs described for the adults of I. auritus [17] .
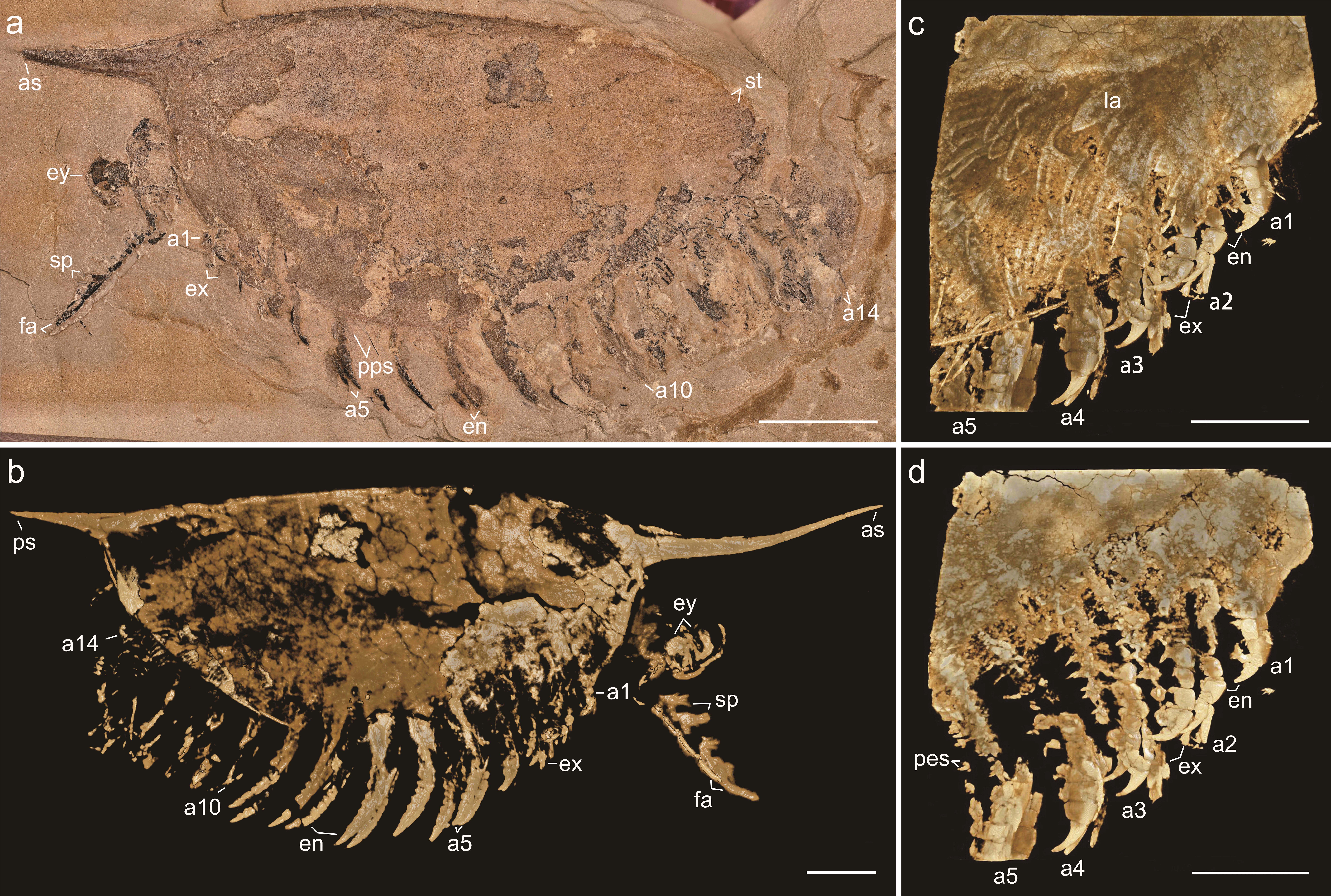
Micro-CT imaging and 3D rendering techniques reveal exceptional details of the pyritized limb morphology in Chengjiang fossil euarthropods that are not accessible through conventional light photography [21, 27, 28]. In I. curvirostratus, all the ventral appendages have a biramous construction, with the first to fourth appendage pairs being shorter and morphologically distinct from the subsequent ones (Fig. 1b–d; Extended Data Fig. 3c, d; Supplementary Videos 1, 2). The endopods have well-defined segmental boundaries expressed as regularly spaced transverse grooves along the proximodistal appendage axis. The fact that the grooves are consistent in their position within and between appendages indicates that they are not fractures in the fossil, nor a result from incomplete pyritization. The number and morphology of the endopod podomeres vary between different appendages. The shorter first to fourth biramous appendage pairs have endopods composed of at least 11 robust podomeres, each with a subtrapezoidal outline, and a strongly curved terminal claw (Figs. 1e–h, and 3c; Supplementary Videos 3–6). Each podomere carries a pair of medially located endites expressed as strong triangular spines along the ventral margin of the main limb axis (Fig. 1f–h; Supplementary Videos 4–6). The exopods of the first to fourth appendage pairs in YKLP 16260 include a slender shaft that is as long as their corresponding endopod (Fig. 1f–h; Supplementary Videos 4–6), and bear several thick paddle-shaped lamellae perpendicular to the main limb axis (Fig. 1c; Extended Data Fig. 3c, d; Supplementary Video 1). The exopod shaft also bears a terminal paddle-shaped lamella with marginal spines on its distal end (Fig. 1f; Supplementary Video 4). The anterior position of the first to fourth appendage pairs and their distinctive morphological specialization suggest that they belong to a functional six-segmented head, which also includes the segments bearing the stalked eyes and the raptorial frontal appendages. The fifth to fourteenth pairs of biramous appendages also show the preservation of fine morphological details, including endopods with at least a dozen well-defined transverse podomere boundaries, and which taper in width distally into a gently curved terminal claw (Fig. 1b, i, j; Extended Data Figs. 2a, 3b). It is likely that the full podomere count is higher, but details of the proximal portion of the appendages cannot be fully resolved. Unlike the cephalic limbs, the fifth to fourteenth pairs of biramous appendages lack endites on each podomere, but feature a pair of elongate delicate multi-articulated spines at the level of the 10th podomere (Fig. 1i, j; Extended Data Fig. 1a; Supplementary Videos 7, 8). The proximal organization of the trunk exopods in I. curvirostratus could not be resolved from the studied material, but the paddle-shaped lamellae are visible on the surface of specimens with well-preserved limbs such as YKLP 16260 (Fig. 1c; Supplementary Video 1).
The limb morphology of Isoxys sp. (Fig. 2; Supplementary Videos 9, 10) complements the findings from I. curvirostratus to produce a comprehensive understanding of the appendicular organization of Isoxys. CFM 00047 features three (rather than four) pairs of smaller biramous appendages, and trunk endopods with more than a dozen podomeres. However, Isoxys sp. demonstrates that the proximal portion of the biramous appendages consists of an undifferentiated subtrapezoidal protopod (Fig. 2e, f; Supplementary Video 12). Isoxys sp. also shows the complete exopod organization in great detail, consisting of an elongate shaft that bears at least a dozen thick and paddle-shaped lamellae that attach perpendicular relative to the main exopod axis (Fig. 2d, f; Supplementary Video 11). The paddle-shaped lamellae imbricate with each other, and lack the fringe of short marginal setae observed in the morphologically similar exopod of Cambrian artiopods [see ref. 28]. Distally, the exopod shaft bears a single paddle-shaped lamella (Fig. 2d–f; Supplementary Videos 11, 12), as also observed in I. curvirostratus (Fig. 1e; Supplementary Video 3). CFM 00047 demonstrates that the imbrication of the paddle-shaped lamellae in the exopod can produce an appearance akin to cuticular folds on the surface of the fossils (Fig. 1l) [15, 25], which has led to previous misinterpretations of the biramous appendage structure.
Appendage differentiation in Isoxys
New material and the use of micro-CT imaging and 3D rendering techniques to detect the iron-enriched non-biomineralized morphology in pyritized Chengjiang fossils reveal new details of the anatomical organization in Isoxys (Fig. 3a–d), with direct implications for its palaeoecological and evolutionary significance. The robust frontal appendages in Isoxys curvirostratus are well suited for a raptorial grasping function following an upwards stroke in front of the carapace (Fig. 3b), which combined with the presence of prominent anterior-facing stalked eyes indicates that this bivalved euarthropod was an active visual predator [10, 12, 14, 26, 29]. The presence of a short unarthrodized trunk concealed within the carapace suggests that this body region was not efficient for swimming on its own, as it lacks the rigid muscle attachment sites needed for propulsion as observed in other Cambrian bivalved forms with cylindrical abdominal tergites [07, 13, 20, 27]. Instead, swimming in Isoxys was most likely achieved by the rhythmic movement of the biramous appendages powered by the paddle-shaped lamellae on the exopods [26, 29] (Fig. 3). The arthropodized trunk endopods would allow walking on the benthos. Whereas the raptorial frontal appendages are well equipped for prey capture thanks to the substantial spinose armature observed in I. curvirostratus (Fig. 1m–o), the following biramous appendages indicate a further degree of functional differentiation. We demonstrate that the four anteriormost biramous appendages of I. curvirostratus are not only shorter, but that their endopods bear robust endites and a strongly curved terminal claw (contra ref. 02, 10, 12, 14, 15, 26) (Fig. 2e–h). The integration of the four biramous appendage pairs into a functionally specialized head region suggest that they were used for processing soft-bodied food items drawn into the anterior space within the carapace – once grasped by the frontal appendages – before consumption. By contrast, the undifferentiated protopod and absence of spinose endites on the trunk biramous appendage pairs – as observed in Isoxys sp. – suggest that they were not used for feeding, but instead were restricted to locomotion for either swimming vertically in the water column [29] or walking on the benthos. The paired slender and multiarticulated spines on the 10th podomere of the trunk endopods are too delicate for food processing; we hypothesize they might have a sensorial function or provide some mechanical support during benthic locomotion. The lack of adaptations for feeding on the trunk biramous appendage pairs also argues against a scavenging or detrivorous diet, as these strategies generally require a dense proximal enditic armature that forms a median food groove for processing organic matter [21, 28]. These findings indicate that the biramous appendages of Isoxys possessed a higher degree of heteronomy and functional differentiation than previously considered [01, 10, 12, 14], and reveal an unexpected complexity in the feeding ecology of pelagic predators in early marine ecosystems [26, 29].
Implications of early euarthropod evolution
Isoxys curvirostratus uniquely combines the presence of fully arthropodized biramous appendages, a morphologically and functionally specialized six-segmented head, and lack of trunk arthrodization (Figs. 3, 4), all of which are critical characters for reconstructing early euarthropod evolution [01, 02, 08, 13]. Notably, I. curvirostratus and Isoxys sp. demonstrate that all the biramous appendages share similarities with those of deuteropods in a more crown-wards phylogenetic position [08]. The multi-podomerous endopods of Isoxys are comparable to those of other Cambrian bivalved euarthropods [17, 20] and fuxianhuiids [06], suggesting that they could reflect the ancestral organization of the earliest arthropodized limbs. Similarly, the slender exopod shaft with paddle-shaped lamellae has recently been recognized in a number of Cambrian artiopods, and suggested as potentially symplesiomorphic for that clade [28]. The Burgess Shale Surusicaris also has three anteriormost differentiated ventral appendages [15], indicating that the presence of a multi-segmented head region is widespread among isoxyids. Critically, an isoxyid-like cephalization pattern has been recently recognized in the early diverging stem-group euarthropod Kylinxia from Chengjiang [02], which strikingly also possesses upwards-facing raptorial frontal appendages and four pairs of smaller biramous appendages in the head. Isoxys and Kylinxia also share the presence of paired telson flaps, although other aspects of their body morphology differ substantially, such as the presence of trunk tergites and absence of a carapace in the latter. Indeed, the lack of trunk arthrodization in Isoxys is comparable with the absence of epidermal dorsal segmentation observed in radiodonts [03, 23, 30], which suggests that Isoxys embodies an earlier step in the evolutionary history of the euarthropod body plan relative to the fully arthrodized Kylinxia.
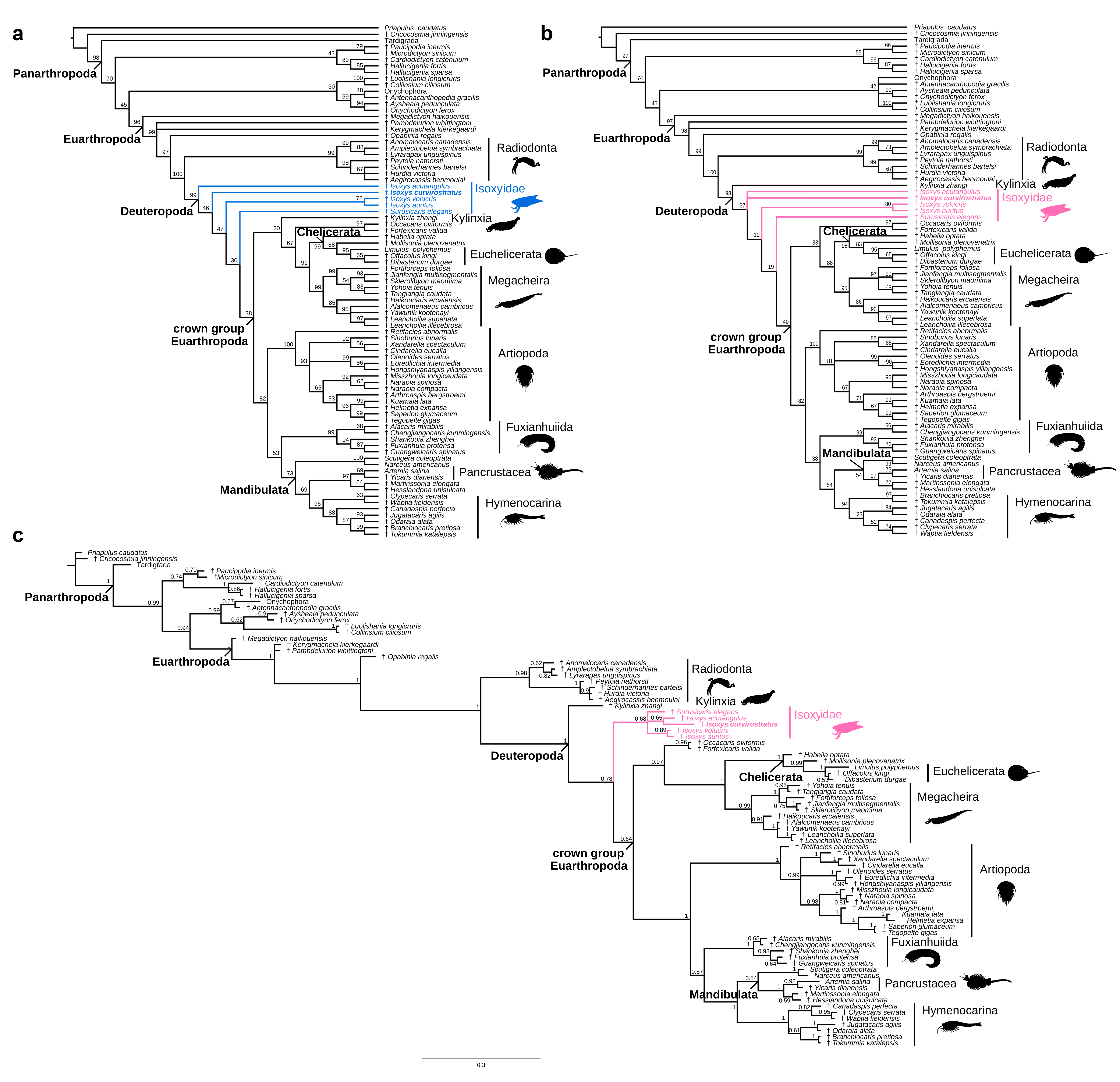
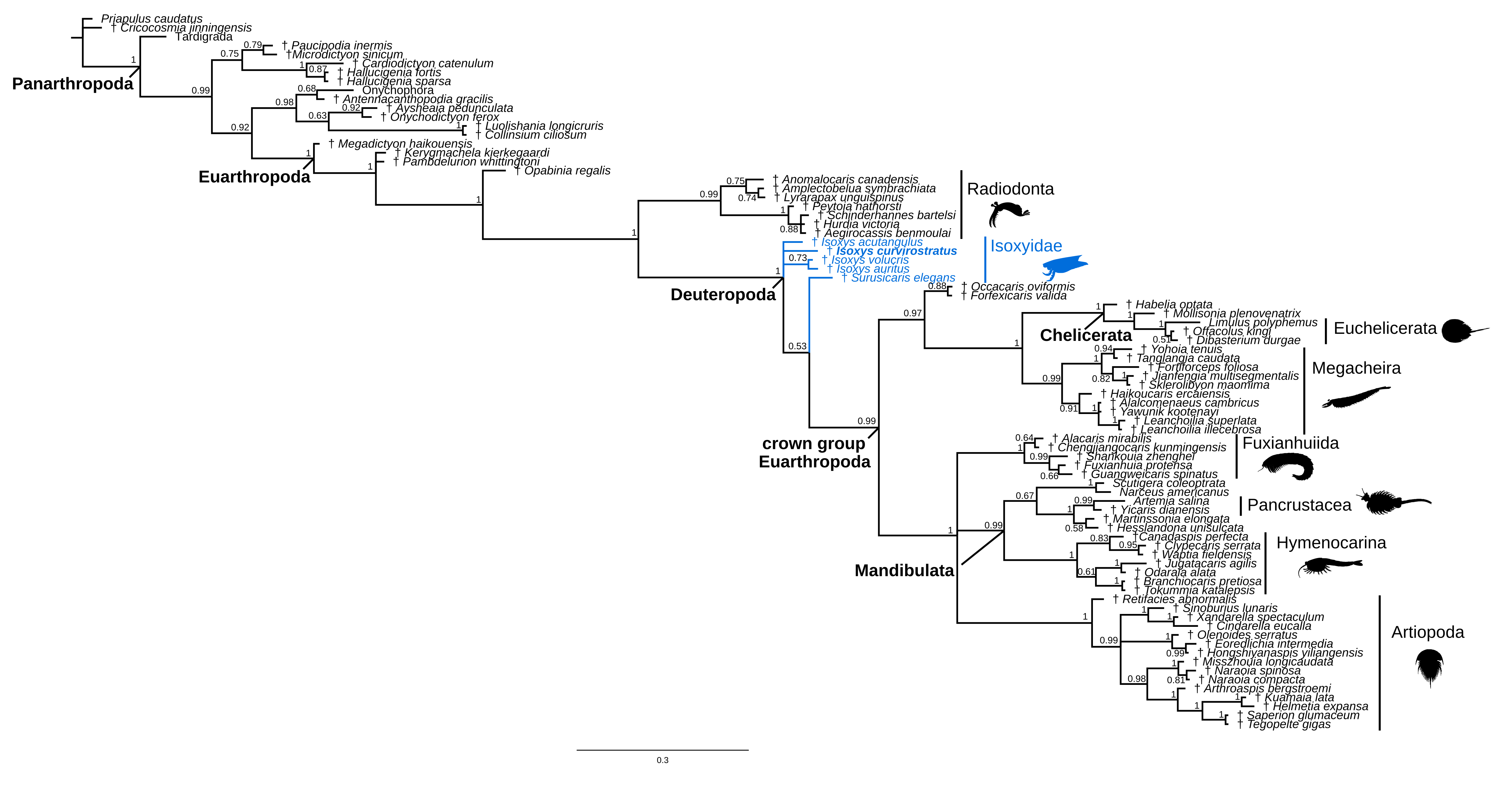
The results of phylogenetic analyses using maximum parsimony and Bayesian inference (Fig. 3e–g; Extended Data Figs. 5–7) to explore the evolutionary implications of our new morphological data provide support for the hypothesis that Isoxys may be the outgroup to other deuteropods, rather than Kylinxia (contra ref. 02). The maximum parsimony analysis with implied weights (Fig. 3e; Extended Data Fig. 5a) resolves isoxyids as a paraphyletic grade and the earliest branching members of Deuteropoda, and Kylinxia as a stem-group chelicerate, while equal weights maintain Kylinxia as the outgroup of Deuteropoda in all parsimonious trees (Fig. 3f; Extended Data Fig. 5b). These results carry different evolutionary implications. Under the Isoxys-first scenario, the arthropodized ventral appendages have a single origin at the base of Deuteropoda, and trunk arthrodization is a synapomorphy of crown-group Euarthropoda (Fig. 3e). The Kylinxia-first scenario also supports a single origin for arthropodized ventral appendages, but requires the repeated evolution of trunk arthrodization in Kylinxia, its loss among isoxyids, and reappearance in crown-group Euarthropoda (Fig. 3f). Thus, Isoxys-first represents the most parsimonious hypothesis for the origin of arthropodization and arthrodization within the euarthropod stem lineage from a character transformation perspective.
Bayesian inference supports Kylinxia as the outgroup of Deuteropoda and the monophyly of isoxyids (Fig. 3g; Extended Data Fig. 5c). This result provides two equally parsimonious scenarios of character evolution: either a single origin for trunk arthrodization in Deuteropoda (evidenced by Kylinxia) with secondary loss among monophyletic isoxyids, or the repeated evolution of trunk arthrodization in Kylinxia and crown-group Euarthropoda. Although Kylinxia as the outgroup has a posterior probability of 0.78 (Fig. 3g; Extended Data Fig. 5c), we use treespace analysis [30] to visualize the topologies retrieved by our phylogenetic reconstruction (n = 2317 trees total), and the degree of uncertainty at this node, information that is omitted when viewing a consensus tree alone. Indeed, 20% (n = 464 trees) of the retrieved topologies support isoxyids as the outgroup, a relationship that occupies a comparable amount of treespace as does the alternative Kylinxia despite being visited fewer times by the Markov chains (Fig. 3h; Extended Data Fig. 6). This implies that isoxyids as the outgroup are plausible given the morphological data available. Similar to the results of experiments by ref. 02, removal of Kylinxia from the morphological dataset completely supports paraphyletic isoxyids as the outgroup of all other deuteropods (Extended Data Fig. 7). In this context, it appears that the unique morphology of Kylinxia may be responsible for character conflict within the euarthropod stem lineage. Ultimately, our results provide evidence that isoxyids are viable candidates as the earliest branching members of Deuteropoda. The reinvigorated understanding of the body organization of Isoxys made possible by new fossil material and micro-CT imaging consolidates their key role in the step-wise evolution of the fundamental exoskeletal characters that define crown-group Euarthropoda.
Online content
Any methods, additional references, Nature Research Reporting summary, source data, supplementary information, peer review information; details of author contributions and competing interests; and statements of date and code availability are available at TBC.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}