Rice phyllomicrobiome samples, metagenome read statistics and diversity-indices
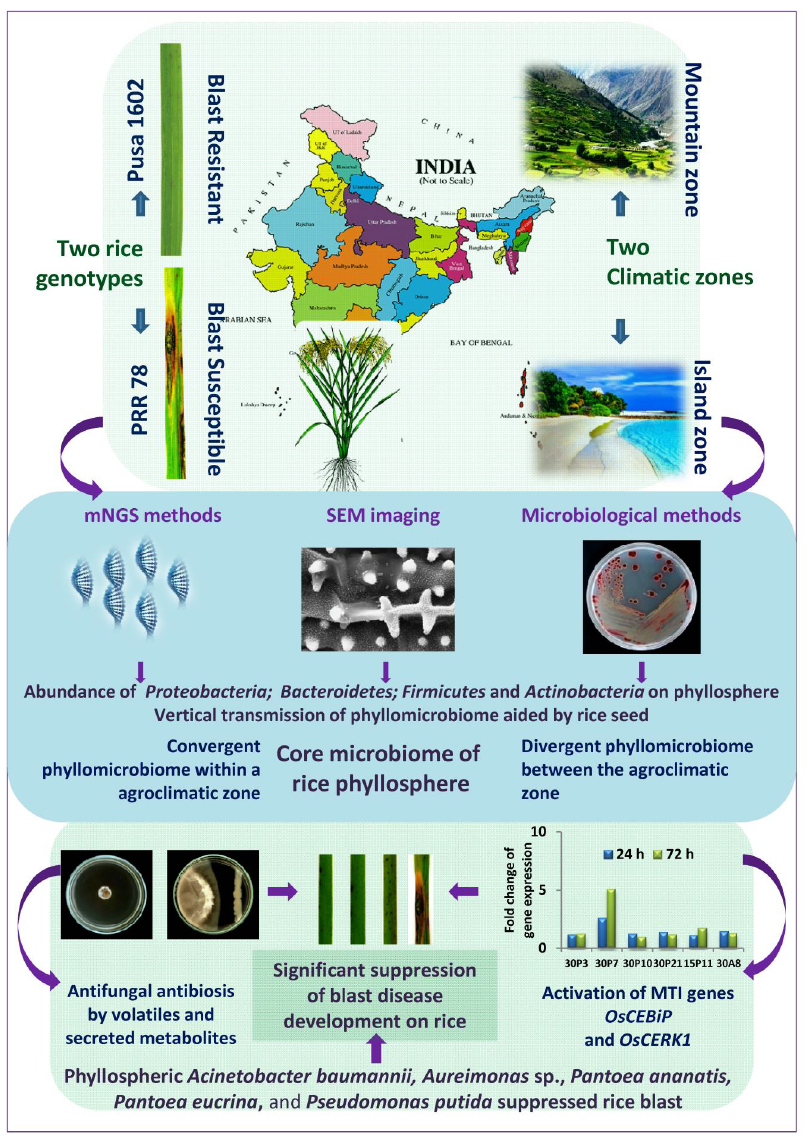
Phyllomicrobiome profiles ofPRR 78 (Blast susceptible) and Pusa 1602 (Blast resistant) grown in two contrasting agro-climatic zones were analysed and decoded by integrated mNGS and microbiological methods (Fig. 1).A total of eight-samples, namely, (i). Palampur-PRR 78-2016 (PRR 78-Plm1 & PRR 78-Plm2); (ii). Palampur-Pusa 1602–2016 (Pusa 1602-Plm1 & Pusa 1602-Plm2); (iii). ANI-PRR 78-2017 (PRR 78-ANI1 & PRR 78-ANI2); (iv). ANI-Pusa 1602–2017 (Pusa 1602-ANI1 & Pusa 1602-ANI2) were generated and subjected to microbiome analysis (Supplementary Table 3). The alpha-diversity indices of phyllosphere-microbial diversity determined using the mNGS data are furnished in Table 1. While the Shannon diversity index ranged from 2.12 to 3.15, the Simpson and Chao1 are in the range of 0.729 to 0.896 and 128.11 to 300.61, respectively. The observed species was in the range of 111–267. The maximum diversity and maximum number of OTUs were found in most of the samples generated from the Island zone (Fig. 2;Table 1).
Table 1
αdiversity of rice phyllomicrobiome representing contrasting agroclimatic zones
|
Location
|
Genotype
|
Samples
|
Shannon
|
Chao1
|
Simpson
|
Observed
|
| |
|
|
Value
|
|
Value
|
SE
|
Value
|
SE
|
Value
|
SE
|
|
Island Zone
|
PRR78
|
PRR78-ANI1-R1
|
2.615
|
-
|
274.65
|
34.14
|
0.875
|
-
|
177
|
-
|
|
Island Zone
|
PRR78
|
PRR78-ANI1-R2
|
2.979
|
-
|
284.00
|
15.95
|
0.896
|
-
|
239
|
-
|
|
Mountain Zone
|
PRR78
|
PRR78-Plm1-R1
|
2.120
|
-
|
128.11
|
8.86
|
0.783
|
-
|
111
|
-
|
|
Mountain Zone
|
PRR78
|
PRR78-Plm2-R2
|
2.356
|
-
|
195.83
|
19.78
|
0.767
|
-
|
148
|
-
|
|
Island Zone
|
Pusa1602
|
Pusa1602-ANI1-R1
|
2.178
|
-
|
263.09
|
22.43
|
0.729
|
-
|
194
|
-
|
|
Island Zone
|
Pusa1602
|
Pusa1602-ANI2-R2
|
2.784
|
-
|
265.88
|
11.74
|
0.843
|
-
|
234
|
-
|
|
Mountain Zone
|
Pusa1602
|
Pusa1602-Plm1-R1
|
2.527
|
-
|
205.24
|
10.36
|
0.815
|
-
|
181
|
-
|
|
Mountain Zone
|
Pusa1602
|
Pusa1602-Plm2-R2
|
3.154
|
-
|
300.61
|
12.14
|
0.881
|
-
|
267
|
-
|
| Microbiome Analyst [37] was utilized for the determination of α-diversity |
PCoA based Bray Curtis and ANoSIM
PCoA of metagenome reads of contrasting rice genotypes, PRR 78, and Pusa 1602 by Bray-Curtis and ANoSIM revealed converging and shared microbiome assemblage on rice genotypes when grown in the same agroclimatic-zone. The same genotype, either PRR 78 or Pusa 1602, showed diverging-microbiome composition when grown in another agroclimatic zone, either Mountain or Island-zone (Fig. 3).
Linear discriminant analysis (LDA) effect size (LEfSe) analysis
The LDA-LEfSescore calculated at 2.0 significance level revealed microbial-biomarker profiles for rice genotypes and agroclimatic zones. The result showed a total of 10 biomarkers for Pusa 1602 and two for PRR 78. Klebsiella and Exiguobacterium were found to be a unique microbial biomarker for PRR 78 while Methylobacterium, Janibacter, Frankia, Macrococcus, Leptolyngbya, Shigella, Pseudacidovorax, Anoxybacillus, and Cellulosimicrobium were predicted to be a biomarker of Pusa 1602. For the geographical location, a total of 15 biomarkers for the mountain zone at Palampur and 16 for the Island zone for Port Blair samples were deciphered.Pantoea, Arthrobacter, Acidovorax, Erwinia, Microbacterium, Shewanella, Acinetobacter, Sphingobacterium, Pseudoalteromonas, Herbaspirillum, Psychrobacter, Candidatus-Koribacter, Mesorhizobium, Variovarax, and Roseateles were found to be a biomarker for mountain zone while, Lysinibacillus, Alkaliphilus, Cylindrospermum, Enterococcus, Bifidobacterium, Arthrospira, Leptolyngbya, Candidatus-Aquiluna, Agromyces, Lactobacillus, Leifsonia, Clostridium, Streptomyces, Bacillus, and Curtobacterium were identified as a biomarker for island zone (SupplementaryFig. 1).
SparCC network of variety and location
Network analysis showed the positive (cooperative) and negative (competitive) interactions within the phyllomicrobiome members on the phyllosphere. In agroclimatic zones and rice genotypes, as many as 68 bacterial genera were predicted to interacting among themselves showing positive and negative interactions on the phyllosphere(Supplementary Table 4; Supplementary Fig. 2). SparCC based network analysis of phyllomicrobiome showed 128 and 127 cooperative interactions on the rice genotypes and agro-climatic zones, respectively; as many as 104 and 108 competitive interactions were also predicted on the genotypes and climatic zones.
Comparative mNGS analysis of contrasting rice genotypes
The bacterial taxa can be considered as a member of “core microbiota” if it is “consistently”associated with all genotypes of a particular species.All other bacterial species may belong to “satellite microbiota” members. Comparative mNGS analysis of rice genotypes revealed the dominance of Proteobacteria, Firmicutes, and Actinobacteria on both the rice genotypes. A total of 11 phyla were found predominated in Pusa 1602 compared to PRR 78; they were Deinococcus-Thermus, Aquificae, Gemmantimonadetes, Chloroflexi, Acidobacteria, Planctomycetes, Verucomicrobia, Actinobacteria, Proteobacteria, Bacteroidetes, and Nitrospirae. On the other hand, only three phyla Firmicutes, Fusobacteria, and Cyanobacteria were found predominated in PRR 78 (Supplementary Fig. 3). Phyllomicrobiome profiles of all taxonomic hierarchies are furnished in Supplementary Fig. 3. Phyllomicrobiome at genus level showed primarily Pantoea followed by Curtobacterium, Methylobacterium, Exiguobacterium, and Bacillus on Pusa 1602; PRR 78showed the dominance of Exiguobacterium followed by Pantoea, Sphingomonas, Curtobacterium, and Arthrobacter (Table 2; Fig. 4).
Table 2
Genotype or climate zone-specific and common bacterial communities associated with phyllosphere of rice
|
Blast Susceptible (PRR78)
|
Prevalence
|
Blast resistant
(Pusa1602)
|
Prevalence
|
Mountain zone
(Palampur)
|
Prevalence
|
Island zone
(Port Blair)
|
Prevalence
|
|
-
|
-
|
Acidovorax
|
0.5
|
Acidovorax
|
1
|
-
|
-
|
|
Arthrobacter
|
0.5
|
Arthrobacter
|
0.5
|
Arthrobacter
|
1
|
-
|
-
|
|
Bacillus
|
0.5
|
Bacillus
|
0.75
|
Bacillus
|
1
|
Bacillus
|
1
|
|
Clavibacter
|
0.25
|
Clavibacter
|
0.25
|
-
|
-
|
Clavibacter
|
0.5
|
|
-
|
-
|
Clostridium
|
0.25
|
-
|
-
|
Clostridium
|
0.25
|
|
-
|
-
|
Cronobacter
|
0.5
|
Cronobacter
|
1
|
Cronobacter
|
0.25
|
|
Curtobacterium
|
0.75
|
Curtobacterium
|
0.75
|
Curtobacterium
|
1
|
Curtobacterium
|
1
|
|
Deinococcus
|
0.25
|
Deinococcus
|
0.5
|
Deinococcus
|
0.5
|
Deinococcus
|
0.5
|
|
Erwinia
|
0.25
|
Erwinia
|
0.25
|
Erwinia
|
0.5
|
-
|
-
|
|
Exiguobacterium
|
1
|
Exiguobacterium
|
1
|
Exiguobacterium
|
0.5
|
Exiguobacterium
|
1
|
|
Hymenobacter
|
0.25
|
Hymenobacter
|
0.5
|
Hymenobacter
|
0.5
|
Hymenobacter
|
0.5
|
|
-
|
-
|
Kineococcus
|
0.5
|
Kineococcus
|
0.5
|
Kineococcus
|
0.25
|
|
Klebsiella
|
1
|
-
|
-
|
Klebsiella
|
0.5
|
Klebsiella
|
0.5
|
|
Methylobacterium
|
0.5
|
Methylobacterium
|
1
|
Methylobacterium
|
0.5
|
Methylobacterium
|
1
|
|
Methylocella
|
0.25
|
-
|
-
|
-
|
-
|
Methylocella
|
0.25
|
|
Microbacterium
|
0.5
|
Microbacterium
|
0.75
|
Microbacterium
|
0.5
|
Microbacterium
|
0.25
|
|
-
|
-
|
Nocardioides
|
0.5
|
Nocardioides
|
0.25
|
-
|
-
|
|
Pantoea
|
1
|
Pantoea
|
1
|
Pantoea
|
0.25
|
Pantoea
|
1
|
|
Pedobacter
|
0.25
|
-
|
-
|
Pedobacter
|
0.25
|
-
|
-
|
|
Pseudomonas
|
0.5
|
-
|
-
|
Pseudomonas
|
0.25
|
-
|
-
|
|
-
|
-
|
Salmonella
|
0.25
|
Salmonella
|
0.25
|
-
|
-
|
|
-
|
-
|
Serratia
|
0.25
|
Serratia
|
0.25
|
-
|
-
|
|
Sphingomonas
|
0.75
|
Sphingomonas
|
0.5
|
Sphingomonas
|
0.25
|
Sphingomonas
|
0.75
|
|
Streptomyces
|
0.25
|
-
|
-
|
-
|
-
|
Streptomyc
|
0.25
|
| Red: Blast susceptible genotype-specific bacterial genera |
| Navy Blue: Blast resistant genotype-specific bacterial genera |
| Green: Mountain zone-specific bacterial genera |
| Dark blue: Island zone-specific bacterial genera |
| Black: Common bacterial genera |
Comparative mNGS analysis of contrasting agroclimatic zones
Comparative mNGS analysis of rice genotypes of two climatic-zones at mountain and island zones revealed the dominance of Proteobacteria, Firmicutes, and Actinobacteria over other phyla on the rice phyllosphere (Supplementary Fig. 4; Supplementary Fig. 5). The comparative mNGS analysis further revealed the dominance of seven phyla each in Port Blair and Palampur samples. Whereas Actinobacteria, Aquificae, Chloroflexi, Cyanobacteria, Nitrospirae, Planctomycetes, and Verucomicrobia were found on the Island zone, themountain zone showed the presence of Acidobacteria, Bacteroidetes Deinococcus-Thermus, Gemmantimonadetes, Firmicutes, Fusobacteria, and Proteobacteria (Supplementary Fig. 4; Supplementary Fig. 5). Unique phyllomicrobiome profiles for mountain and island agroclimatic zones showing taxonomic hierarchies such as class, order, and family are presented in Supplementary Fig. 4 and Supplementary Fig. 5.The genera-level comparative microbial profile revealed the predominance of Bacillus, Curtobacterium, Exiguobacterium, Pantoea, and Sphingomonas at the Island zone while the rice phyllosphere in the mountain zone was dominated by Arthrobacter, Exiguobacterium, Methylobacterium, and Pantoea over other bacterial genera (Table 2; Fig. 4; Fig. 5; Supplementary Fig. 6).
Core microbiome analysis
Core-microbiome at the genus level was analyzed for rice genotypes as well as for the agroclimatic zones. Core microbiome of blast susceptible genotype, PRR 78 was found consisting of 17 bacterial genera with a maximum prevalence of Pantoea, Klebsiella, and Exiguobacterium. Blast resistant genotype Pusa 1602 showed core microbiota composed of 19 genera with the maximum prevalence of Pantoea, Methylobacterium, and Exiguobacterium. For agroclimatic zones, the core phyllomicrobiome at the mountain zone was found comprising of 20 genera with the high representation of Pantoea, Microbacterium Exiguobacterium, and Arthrobacter. Similarly, the core phyllomicrobiome at the Island zone displayed 16 genera with the maximum prevalence of Pantoea, Methylobacterium, Exiguobacterium, Curtobacterium, and Bacillus.
Overall, the core phyllomicrobiome of rice deduced from all sets of samples revealed 26 bacterial genera with the maximum prevalence of PantoeaandExiguobacterium. The other member of rice core phyllomicrobiome were Methylobacterium, Curtobacterium, Sphingomonas, Microbacterium, Bacillus, Klebsiella, Arthrobacter, Hymenobacter, Deinococcus, Pseudomonas, Nocardioides, Kineococcus, Erwinia, Cronobacter, Clavibacter, Acidovorax, Streptomyces, Serratia, Salmonella, Pedobacter, Methylocella, and Clostridium (Table 3).
Table 3
Core phyllomicrobiome of rice genotypes grown in two contrasting climate zones
|
Core phyllomicrobiome
|
Prevalence
|
|
Acidovorax
|
0.25
|
|
Arthrobacter
|
0.5
|
|
Bacillus
|
0.625
|
|
Clavibacter
|
0.25
|
|
Clostridium
|
0.125
|
|
Cronobacter
|
0.25
|
|
Curtobacterium
|
0.75
|
|
Deinococcus
|
0.375
|
|
Erwinia
|
0.25
|
|
Exiguobacterium
|
1.0
|
|
Hymenobacter
|
0.375
|
|
Kineococcus
|
0.25
|
|
Klebsiella
|
0.5
|
|
Methylobacterium
|
0.75
|
|
Methylocella
|
0.125
|
|
Microbacterium
|
0.625
|
|
Nocardioides
|
0.25
|
|
Pantoea
|
1.0
|
|
Pedobacter
|
0.125
|
|
Pseudomonas
|
0.25
|
|
Salmonella
|
0.125
|
|
Serratia
|
0.125
|
|
Sphingomonas
|
0.625
|
|
Streptomyces
|
0.125
|
| Microbiome Analyst [37] was utilized for the determination of core phyllomicrobiome |
Scanning Electron Microscopic imaging of phyllomicrobiome
The SEM imaging of rice phyllomicrobiome revealed the physical presence of bacterial cells aggregates of 5–8 cells, and unevenly distributed solitary bacterial-cells on the phyllosphere of rice genotypes. The Eukaryotic cells and hyphal fragments were also found scattered among the prokaryotic cells (Fig. 6).
Culturable phyllomicrobiome analysis
Enumeration, characterization, and identification of rice phyllomicrobiome associated bacterial communities: Susceptible genotype (3.127–4.313 CFU g− 1) recorded the higher epiphytic bacterial population as compared to resistant genotypes (2.945–3.317CFU g− 1) in both locations (Supplementary Table 5; Supplementary Table 6). A total of 78 distinct morphotypes of cultured bacterial communities were isolated from both locations. A relatively more bacterial population was found on 30 days old phyllosphere (45 morphotypes) as compared to 15 days (33 morphotypes). The results of diversity indices indicated that the blast susceptible genotype and 30 days old phyllosphere recorded significantly more bacterial diversity than the resistant genotype and 15 days old phyllosphere. The Shannon diversity index ranged from 1.12 to 1.8 for all the cultured phyllosphere microbiome. The diversity indices of epiphytic bacteria isolate colonized rice phyllosphere representing three locations are presented in Table 4. BOX-PCRDNA fingerprinting of all 78 morphotypes culminated in 59 distinct BOX Amplicon Groups. At least in one BOX-amplicon group, the amplicon profiles were found perfectly identical for isolates OsEp-Plm-15P4; OsEp-Plm-15P8; OsEp-Plm-15P9; OsEp-Plm-15P10; OsEp-Plm-15P13; OsEp-Plm-15P15 from mountain zone, and for isolates, OsEp-AN-15A10; OsEp-AN-15A11; OsEp-AN-15A17; OsEp-AN-15A18 representing island zone. One of each isolates, OsEp-Plm-15P9 and OsEp-AN-15A10, representing the mountain zone and island zone, respectively were selected for further work (Supplementary Fig. 7). Isolates with identical amplicon profiles were considered duplicates. 16S rRNA gene sequence-based database searches for isolated bacterial species revealed the high-frequency occurrence of Acidovorax (3), Acinetobacter (6), Aureimonas (2), Curtobacterium (5), Enterobacter (6), Exiguobacterium (4), Microbacterium (2), Pantoea (16), Pseudomonas(5) and Sphingomonas (7) on rice phyllosphere (Supplementary Fig. 8; Supplementary Table 7). Six bacterial isolates from the mountain zone and four from the island zone (represented by OsEp-Plm-15P9 and OsEp-AN-15A10) which shared all BOX PCR amplicons (genetically identical isolates) were identified as Pantoea ananatis.
Table 4
Diversity indices of cultured-phyllosphere bacterial communities representing three geographical locations
|
Rice phyllosphere
|
Parameters
|
*Age of plantlets
|
|
15
|
30
|
|
Pusa1602
|
PRR78
|
Pusa1602
|
PRR78
|
|
Palampur
|
Shannon Wiener diversity index
|
1.30
|
1.80
|
1.80
|
1.58
|
|
Species richness
|
10.0
|
10.0
|
15.0
|
17.0
|
|
Port Blair
|
Shannon Wiener diversity index
|
1.12
|
1.40
|
1.40
|
1.34
|
|
Species richness
|
12.0
|
9.0
|
17.0
|
17.0
|
| * Days Post Transplanting |
Microbiological validation of phyllomicrobiome profile and isolation of core microbiome
A total of 59 bacterial species belonging to 14 diverse bacterial genera such as Acidovorax, Acinetobacter, Agrobacterium, Aureimonas, Curtobacterium, Enterobacter, Enterococcus, Erwinia, Exiguobacterium, Microbacterium, Micrococcus, Pantoea, Pseudomonas, and Sphingomonas were cultured, isolated, and preserved from rice phyllomicrobiome (Supplementary Fig. 9a-9m). All cultured bacterial flora were also found among the mapped reads in the mNGS data. Further, comparative analysis of phyllomicrobiomeof rice samples confirmed the consistent association of Acinetobacter, Curtobacterium, Enterobacter, Exiguobacterium, Pantoea, Pseudomonas, and Sphingomonas in Mountain and Island agroclimatic zones in both the mNGS and microbiological approaches (Data not shown). Bacterial genera suchas Acinetobacter, Curtobacterium, Enterobacter, Exiguobacterium, Pantoea, Pseudomonas, and Sphingomonas were consistently associated with both the genotypesin all samples(data not shown).
Activity screening for identification of functional core-phyllomicrobiome
Screening for antifungal activity: Dual plate confrontation assay showed inhibition of mycelial growth of M. oryzaeby both volatiles and secreted metabolites produced by bacterial species. Among the 59 bacteria evaluated, 14 phyllosphere-associated bacterial isolates (23.7%) displayed over 40.0 % inhibition of mycelial growth by their secreted metabolites (Table 5; Supplementary Fig. 10). The antagonistic bacterial isolates represented species such as Acinetobacter baumannii; Acinetobacter soli; Erwinia tasmaniensis; Exiguobacterium indicum; Pantoea agglomerans; Pantoea ananatis; Pantoea dispersa; Pantoea eucrina; and Pseudomonas oryzihabitans. Similarly, a total of 15 of them (25.4 %) inhibited the growth of M. oryzae completely by airborne bacterial volatile organic compounds (BVCs) (Table 5; Supplementary Fig. 11). The antifungal volatile releasing bacterial isolates represented the species such as Acinetobacter baumannii; Acinetobacter soli; Aureimonas sp.; Pantoea agglomerans; Pantoea ananatis; Pantoea dispersa; Pantoea eucrina; Pseudomonas parafulva, Pseudomonas putida; andPseudomonas oryzihabitans. Further,the BVCs of five bacterial isolates were found to show fungicidal activity while the remaining 10 were fungistatic on M. oryzae(SupplementaryTable 6; Supplementary Fig. 12).
Table 5
Antifungal antagonistic activity displayed by bacterial communities associated with phyllomicrobiome on Magnaporthe oryzae
|
Genus
|
Bacterial isolate
(*Closet Match)
|
*Sequence length
(bp)
|
*GenBank Accession
|
Geographical Zone
|
Mycelial Inhibition (%)
|
|
BVC
|
SC
|
|
Acidovorax
|
Acidovorax avenae OsEp-Plm-30P1
|
1433
|
MT367817
|
Mountain zone
|
34.3
|
3.7
|
|
Acidovorax avenae OsEp-Plm-30P23
|
1378
|
MT367833
|
Mountain zone
|
27.9
|
12.0
|
|
Acidovorax avenae OsEp-Plm-30P6
|
1396
|
MT367820
|
Mountain zone
|
29.3
|
4.6
|
|
Acinetobacter
|
Acinetobacter baumannii OsEp-Plm-30P11
|
1430
|
MT367824
|
Mountain zone
|
100.0
|
39.8
|
|
Acinetobacter baumannii OsEp-Plm-30P17
|
1401
|
MT367827
|
Mountain zone
|
100.0
|
50.9
|
|
Acinetobacter junii OsEp-AN-30A17
|
1386
|
MT367859
|
Island zone
|
52.9
|
7.4
|
|
Acinetobacter soli OsEp-Plm-30P2
|
1419
|
MT394056
|
Mountain zone
|
32.9
|
39.8
|
|
Acinetobacter soli OsEp-Plm-30P4
|
1429
|
MT367819
|
Mountain zone
|
100.0
|
42.6
|
|
Acinetobacter soliOsEp-Plm-30P22
|
1417
|
MT367832
|
Mountain zone
|
32.1
|
34.3
|
|
Agrobacterium
|
Agrobacterium larrymoorei OsEp-Plm-30P19
|
1359
|
MT367829
|
Mountain zone
|
46.4
|
5.6
|
|
Aureimonas
|
Aureimonas phyllosphaerae OsEp-AN-30A11
|
1390
|
MT367855
|
Island zone
|
33.6
|
6.5
|
|
Aureimonas sp.OsEp-Plm-30P7
|
1369
|
MT367821
|
Mountain zone
|
100.0
|
4.6
|
|
Curtobacterium
|
Curtobacterium albidum OsEp-Plm-15P1
|
1391
|
MT367807
|
Mountain zone
|
32.1
|
1.9
|
|
Curtobacterium albidum OsEp-Plm-30P20
|
1401
|
MT367830
|
Mountain zone
|
57.9
|
7.4
|
|
Curtobacterium citreum OsEp-AN-30A1
|
1395
|
MT367846
|
Island zone
|
40.0
|
10.2
|
|
Curtobacterium luteum OsEp-Plm-30P9
|
1393
|
MT367822
|
Mountain zone
|
39.3
|
13.9
|
|
Curtobacterium luteum OsEp-Plm-15P7
|
1390
|
MT367812
|
Mountain zone
|
60.0
|
4.6
|
|
Enterobacter
|
Enterobacter asburiae OsEp-AN-30A22
|
1406
|
MT367864
|
Island zone
|
23.6
|
6.5
|
|
Enterobacter asburiae OsEp-Plm-30P16
|
1410
|
MT367826
|
Mountain zone
|
41.4
|
35.2
|
|
Enterobacter cloacae OsEp-AN-15A7
|
1409
|
MT367840
|
Island zone
|
0.0
|
7.4
|
|
Enterobacter cloacae OsEp-Plm-30P18
|
1425
|
MT367828
|
Mountain zone
|
18.6
|
23.2
|
|
Enterobacter mori OsEp-AN-30A20
|
1409
|
MT367862
|
Island zone
|
25.7
|
9.3
|
|
Enterobacter sichuanensis OsEp-AN-15A12
|
1404
|
MT367844
|
Island zone
|
41.4
|
5.6
|
|
Erwinia
|
Erwinia tasmaniensis OsEp-AN-15A5
|
1412
|
MT367838
|
Island zone
|
56.4
|
54.6
|
|
Exiguobacterium
|
Exiguobacterium acetylicum OsEp-Plm-15P3
|
1438
|
MT367809
|
Mountain zone
|
54.3
|
1.9
|
|
Exiguobacterium indicum OsEp-AN-30A4
|
1413
|
MT367849
|
Island zone
|
63.6
|
46.3
|
|
Exiguobacterium indicum OsEp-AN-30A6
|
1430
|
MT367851
|
Island zone
|
32.1
|
14.8
|
|
Exiguobacterium indicum OsEp-Plm-30P14
|
1431
|
MT367825
|
Mountain zone
|
24.3
|
3.7
|
|
Microbacterium
|
Microbacterium sp. OsEp-AN-15A2
|
1387
|
MT367835
|
Island zone
|
0.0
|
13.9
|
|
Microbacterium testaceum OsEp-AN-30A2
|
1409
|
MT367847
|
Island zone
|
47.1
|
38.9
|
|
Micrococcus
|
Micrococcus luteus OsEp-AN-15A1
|
1400
|
MT367834
|
Island zone
|
0.0
|
12.0
|
|
Pantoea
|
Pantoea agglomerans OsEp-AN-15A8
|
1418
|
MT367841
|
Island zone
|
69.3
|
7.4
|
|
Pantoea agglomerans OsEp-AN-30A14
|
1408
|
MT367857
|
Island zone
|
100.0
|
42.6
|
|
Pantoea agglomerans OsEp-AN-30A21
|
1413
|
MT367863
|
Island zone
|
40.0
|
10.2
|
|
Pantoea ananatis OsEp-AN-15A10
|
1401
|
MT367843
|
Island zone
|
81.4
|
50.0
|
|
Pantoea ananatis OsEp-AN-30A19
|
1408
|
MT367861
|
Island zone
|
30.7
|
7.4
|
|
Pantoea ananatis OsEp-AN-30A5
|
1402
|
MT367850
|
Island zone
|
100.0
|
4.6
|
|
Pantoea ananatis OsEp-AN-30A8
|
1403
|
MT367852
|
Island zone
|
100.0
|
7.4
|
|
Pantoea ananatis OsEp-Plm-15P9
|
1410
|
MT367813
|
Mountain zone
|
100.0
|
34.3
|
|
Pantoea ananatis OsEp-Plm-30P21
|
1405
|
MT367831
|
Mountain zone
|
100.0
|
50.0
|
|
Pantoea ananatis OsEp-Plm-30P3
|
1419
|
MT367818
|
Mountain zone
|
74.3
|
50.0
|
|
Pantoea dispersa OsEp-AN-30A18
|
1412
|
MT367860
|
Island zone
|
100.0
|
48.2
|
|
Pantoea eucrina OsEp-AN-15A4
|
1409
|
MT367837
|
Island zone
|
100.0
|
50.0
|
|
Pantoea eucrina OsEp-Plm-15P14
|
1421
|
MT367816
|
Mountain zone
|
100.0
|
52.8
|
|
Pantoea eucrina OsEp-Plm-30P10
|
1414
|
MT367823
|
Mountain zone
|
100.0
|
47.2
|
|
Pantoea sp. OsEp-AN-15A15
|
1400
|
MT367845
|
Island zone
|
57.1
|
49.1
|
|
Pantoea sp. OsEp-AN-15A9
|
1402
|
MT367842
|
Island zone
|
0.0
|
3.7
|
|
Pseudomonas
|
Pseudomonas oryzihabitans OsEp-Plm-15P6
|
1398
|
MT367811
|
Mountain zone
|
56.4
|
51.9
|
|
Pseudomonas parafulva OsEp-Plm-15P12
|
1407
|
MT367815
|
Mountain zone
|
100.0
|
38.9
|
|
Pseudomonas psychrotolerans OsEp-AN-15A6
|
1383
|
MT367839
|
Island zone
|
38.6
|
36.1
|
|
Pseudomonas psychrotolerans OsEp-AN-30A13
|
1396
|
MT367856
|
Island zone
|
57.1
|
26.9
|
|
Pseudomonas putida OsEp-Plm-15P11
|
1401
|
MT367814
|
Mountain zone
|
100.0
|
19.4
|
|
Sphingomonas
|
Sphingomonas paucimobilis OsEp-AN-15A3
|
1390
|
MT367836
|
Island zone
|
4.3
|
13.0
|
|
Sphingomonas paucimobilis OsEp-AN-30A9
|
1377
|
MT367853
|
Island zone
|
61.4
|
22.2
|
|
Sphingomonas pseudosanguinis OsEp-AN-30A10
|
1378
|
MT367854
|
Island zone
|
59.3
|
24.1
|
|
Sphingomonas pseudosanguinis OsEp-Plm-15P2
|
1389
|
MT367808
|
Mountain zone
|
79.3
|
15.7
|
|
Sphingomonas sp. OsEp-AN-30A15
|
1362
|
MT367858
|
Island zone
|
60.7
|
4.6
|
|
Sphingomonas sp. OsEp-Plm-15P5
|
1378
|
MT367810
|
Mountain zone
|
58.6
|
3.7
|
|
Sphingomonas yabuuchiae OsEp-AN-30A3
|
1362
|
MT367848
|
Island zone
|
35.7
|
6.5
|
| |
Mock
|
|
|
Both zones
|
0.0
|
0.0
|
| *16S rRNA gene sequences as accessed in https://blast.ncbi.nlm.nih.gov/Blast.cgi during December 2020 |
Screening for blast disease suppression: Blast susceptible rice cultivar, Pusa Basmati 1, was used for evaluating the anti-blast activity of rice phyllomicrobiome associated bacterial communities on blast disease incited by M. oryzae. A total of 20bacterial strains were selected based on in vitro inhibition of M. oryzae. The isolates represented bacterial genera suchas Pantoea (12 strains), Pseudomonas (2), Acinetobacter (3), Aureimonas (1), Erwinia (1), and Exiguobacterium (1). Rice seeds germinated in the presence of bacterial cells (2×107 CFU mL− 1) were allowed to grow in a climate-controlled green-house and challenged withM. oryzae.Before pathogen challenge inoculation, a booster dose of bacterial cell suspension was sprayed onto the leaf lamina. Blast incidence and severity were scored as per the blast score chart recommended by Mackill and Bonman [43]. Most of the bacterial isolates were found to reduce the blast disease development in the plants of the susceptible rice cultivar. Maximum reduction in disease severity was shown by Pantoea ananatis OsEp-Plm-30P3 (74.3%), Pantoea ananatis OsEp-Plm-30P21 (74.2%), Pantoea ananatis OsEp-AN-30A8 (73.0%.), Aureimonas sp.OsEp-Plm-30P7 (73.0%), Pantoea eucrina OsEp-Plm-30P10 (71.5%),Pseudomonas putida OsEp-Plm-15P11 (51.8 %), Pantoea ananatis OsEp-Plm-15P9 (49.7%), and Acinetobacter baumannii OsEp-Plm-30P11 (47.3%) (Table 6; Fig. 7; Supplementary Fig. 13).
Table 6
Blast suppressive potential showed by phyllosphere bacterial genera on rice
|
Genus
|
Bacterial isolates
|
*Blast disease suppression
|
|
*Severity Score
|
*Severity Reduction (%)
|
|
Acinetobacter
|
Acinetobacter baumannii OsEp-Plm-30P11
|
26.8
|
47.3
|
|
Acinetobacter baumannii OsEp-Plm-30P17
|
28.6
|
43.7
|
|
Acinetobacter soli OsEp-Plm-30P4
|
33.3
|
34.5
|
|
Aureimonas
|
Aureimonas sp. OsEp-Plm-30P7
|
13.7
|
73.0
|
|
Erwinia
|
Erwinia tasmaniensis OsEp-AN-15A5
|
33.5
|
34.2
|
|
Exiguobacterium
|
Exiguobacterium indicum OsEp-AN-30A4
|
33.0
|
35.0
|
|
Pantoea
|
Pantoea agglomerans OsEp-AN-30A14
|
29.7
|
41.5
|
|
Pantoea ananatis OsEp-Plm-30P3
|
13.1
|
74.3
|
|
Pantoea ananatis OsEp-Plm-30P21
|
13.1
|
74.2
|
|
Pantoea ananatis OsEp-AN-30A8
|
13.7
|
73.0
|
|
Pantoea ananatis OsEp-Plm-15P9
|
25.6
|
49.7
|
|
Pantoea ananatis OsEp-AN-15A10
|
27.2
|
46.6
|
|
Pantoea ananatis OsEp-AN-30A5
|
30.4
|
40.2
|
|
Pantoea dispersa OsEp-AN-30A18
|
31.2
|
38.5
|
|
Pantoea eucrina OsEp-Plm-30P10
|
14.5
|
71.5
|
|
Pantoea eucrina OsEp-AN-15A4
|
27.1
|
46.7
|
|
Pantoea eucrina OsEp-Plm-15P14
|
28.0
|
45.0
|
|
Pantoea sp. OsEp-AN-15A15
|
27.2
|
46.5
|
|
Pseudomonas
|
Pseudomonas parafulva OsEp-Plm-15P12
|
32.5
|
36.2
|
|
Pseudomonas putida OsEp-Plm-15P11
|
24.5
|
51.8
|
|
Pathogen-Check
|
Control
|
50.8
|
0.0
|
|
Fungicide-Check
|
Tricyclazole control
|
8.33
|
83.6
|
| *Average of three repeat trials each with five replications |
Phyllosphere bacteria-induced expression of defense genes in rice
Seven candidate plant defense genes i.e. OsCEBiP, OsCERK1, OsPAD4, OsNPR1, OsPDF2.2, OsFMO1, and OsPR1.1 showed marginal to a high level of expression in phyllobacterised rice seedlings as compared to the reference gene, OsActin. Interestingly, all six phyllosphere bacterial speciessuch asPantoea ananatis OsEp-Plm-30P3; Aureimonas sp. OsEp-Plm-30P7; Pantoea eucrina OsEp-Plm-30P10; Pantoea ananatis OsEp-Plm-30P21; Pseudomonas putida OsEp-Plm-15P11 and Pantoea ananatis OsEp-AN-30A8 triggered consistent over-expression of OsCEBiP in rice seedlings. Significant induction of OsCEBiP, OsCERK1, andOsPAD4was observed in rice seedlings sprayed with Pantoea or Aureimonas.Strikingly, Aureimonas sp. OsEp-Plm-30P7 showed significant and sustained over-expression of OsCEBiPin 24, 48, and 72 hpi. The epiphytic bacteria-inoculation mediated activation of defense genes was more pronounced during early time points peaking at 48 hpi with a sharp drop at 72 h of bacterial interaction (Fig. 8; Supplementary Fig. 14; Supplementary Table 9).



{kind=link}