Determining the impact that colonisation by GI helminth parasites exerts on the composition of the gut microbial flora of their vertebrate hosts is key to the identification of populations of bacteria that may play active roles in immune- and/or pathophysiological mechanisms that contribute to the outcome of infection and disease; in turn, this new knowledge could be exploited for the development of novel treatment and/or control strategies against parasitic diseases based on the manipulation of the gut microbiota. To this end, we characterised the qualitative and quantitative fluctuations in gut microbiota composition of sheep experimentally infected with a socioeconomically important GI nematode, T. circumcincta, and investigated the relative contribution of vaccine-induced host immunity to the observed changes in the proportions of populations of resident bacteria with potential roles in host-parasite interactions. Furthermore, a novel high-throughput cytometry method, based on immunofluorescence labelling coupled to computational image analysis and cell scoring [37], was set up for the study of T cell populations in the ovine abomasum, and their correlation with parasite load and the abundance of certain bacteria in the faeces of a subgroup of vaccinated and unvaccinated animals.
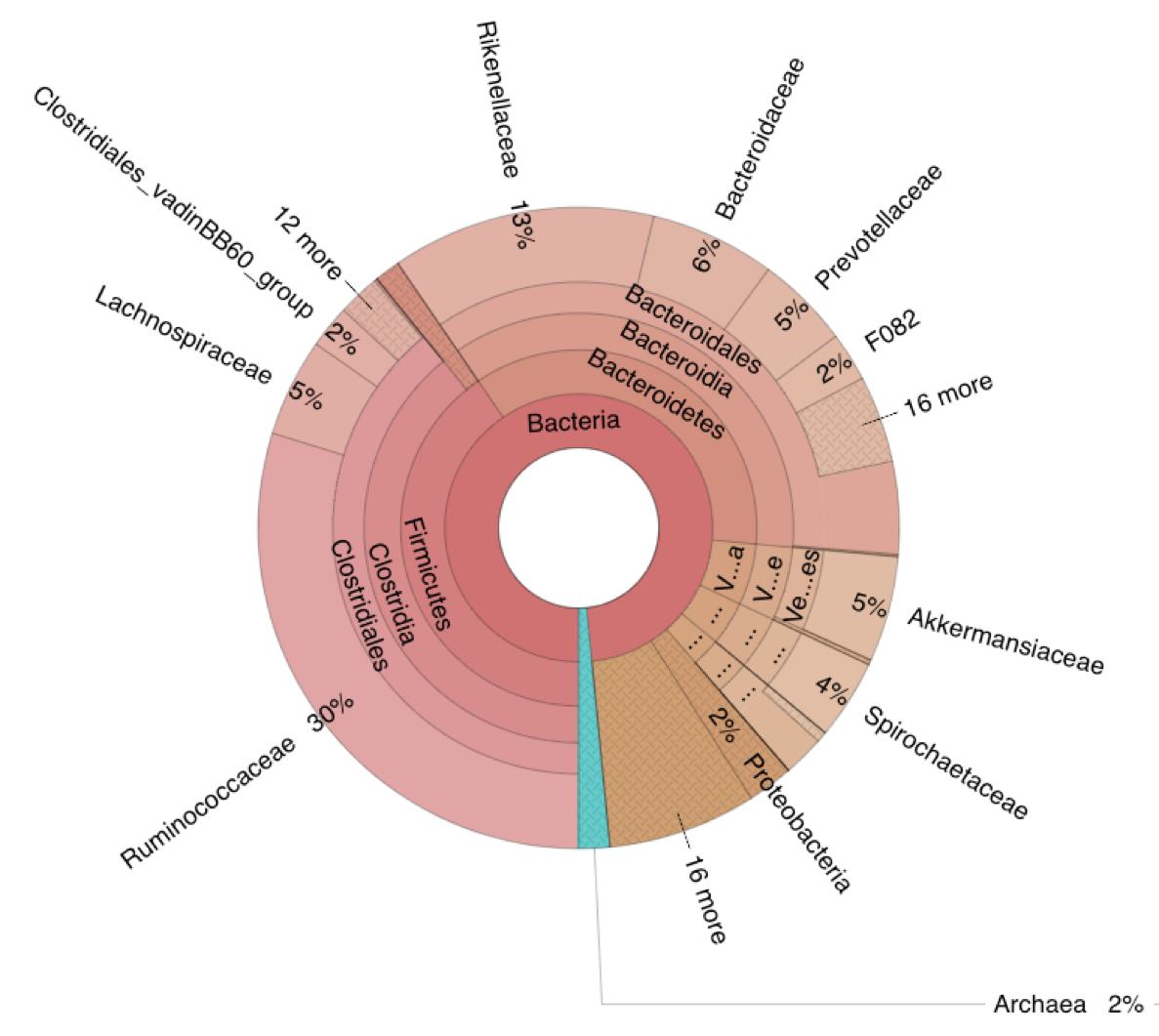
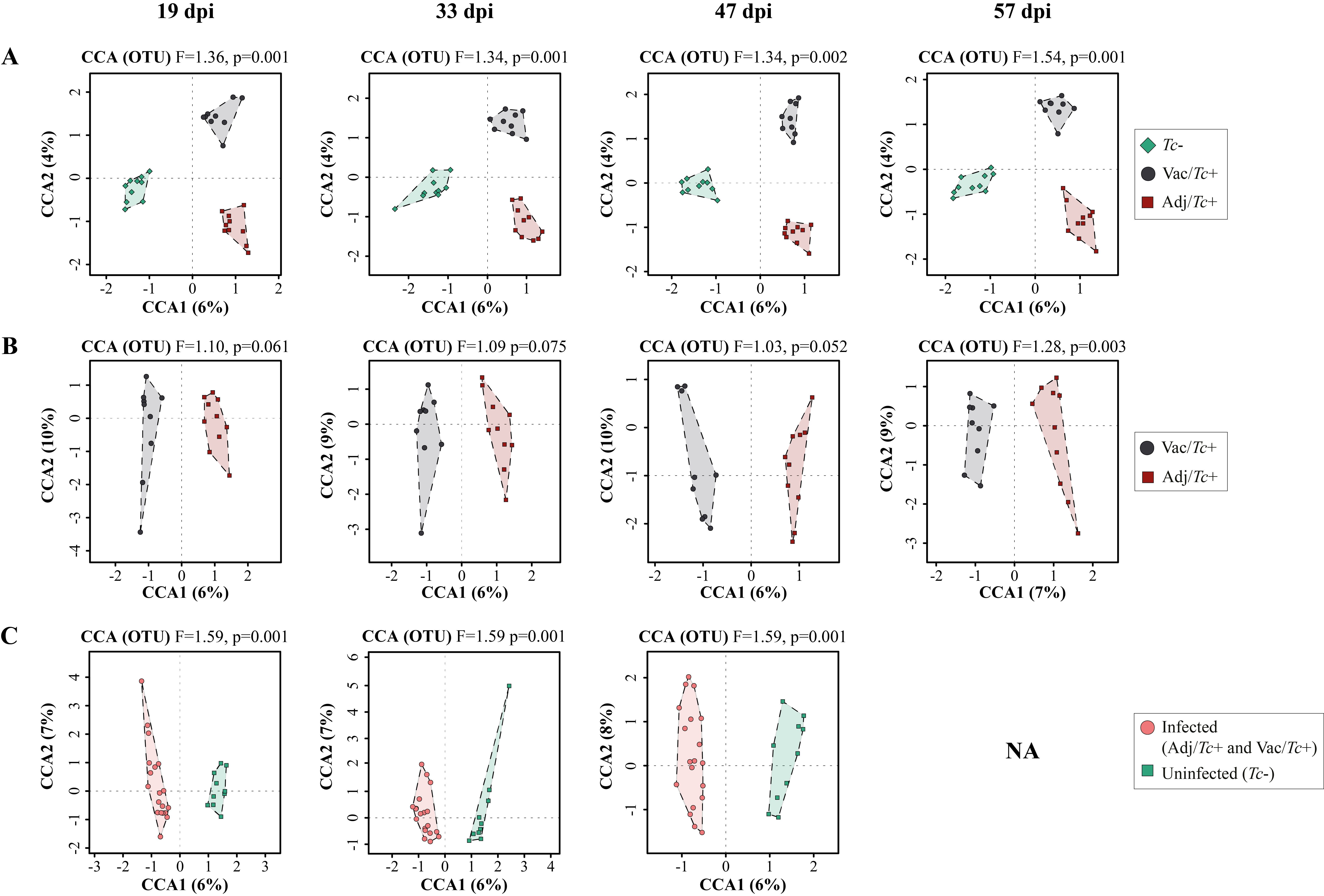
The overall gut microbial profiles of sheep enrolled in this study (i.e. Adj/Tc+, Vac/Tc+ and Tc-)) was consistent with previous descriptions of the ovine gut microbiota [38–39]. Nonetheless, experimental infection with T. circumcincta induced significant changes in the taxonomic profiles of sheep gut communities that occurred irrespective of previous vaccination. Quantitative and qualitative alterations in gut microbial populations in response to GI helminth infections have been widely reported in both humans and veterinary species (reviewed by [6,40]) including in ruminants infected with abomasal nematodes [41–43]. Indeed, whilst our study used faecal microbial composition as a proxy of bacterial populations inhabiting other compartments of the sheep GI tract (cf. 44; reviewed by 45), selected alterations observed herein are consistent with previous descriptions of qualitative and quantitative changes of bacterial populations inhabiting the abomasa of ruminants infected with GI nematodes [41–43], and thus provide support to the validity of our findings. Taken together, these data confirm the major role of helminth colonisation in gut microbiome re-modelling, and that experimental vaccination prior to parasite challenge resulted in further alterations of the gut microbial profiles of Vac/Tc+ sheep at the end of the study period, which were likely to result from the effect of vaccination on worm survival. Indeed, vaccination alone did not result in significant changes in faecal microbial composition at the end of the immunisation period, with the exception of samples collected at the end of the immunisation period, for which marginally significant differences were detected by Adonis between the two sample groups, that were however not supported by CCA and RDA. Nevertheless, the absence of a vaccine-only group (a choice guided by the 3R principles) prevents from speculating on the occurrence of a complex three-way network of interactions between the parasite, gut microbiome and host immune system.
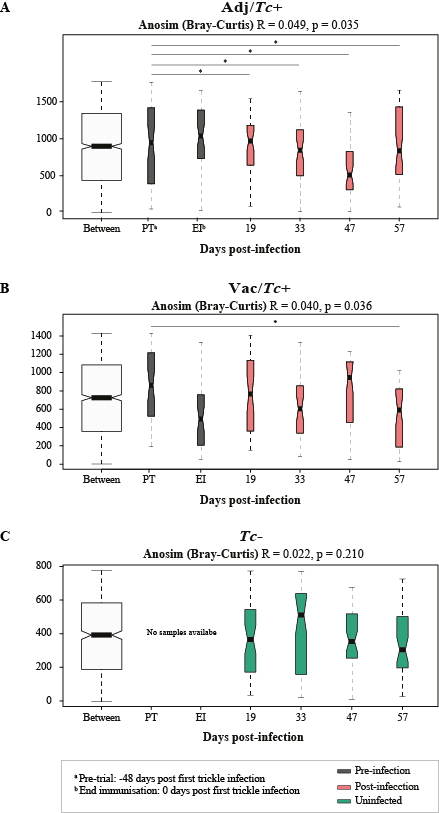
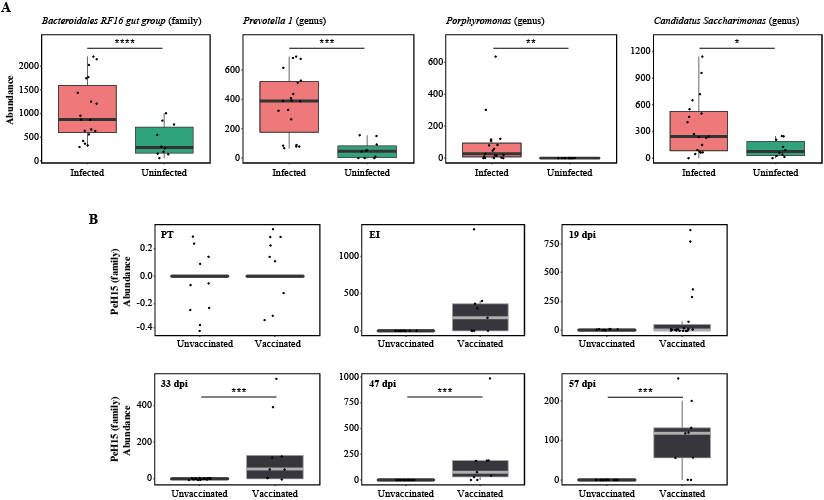
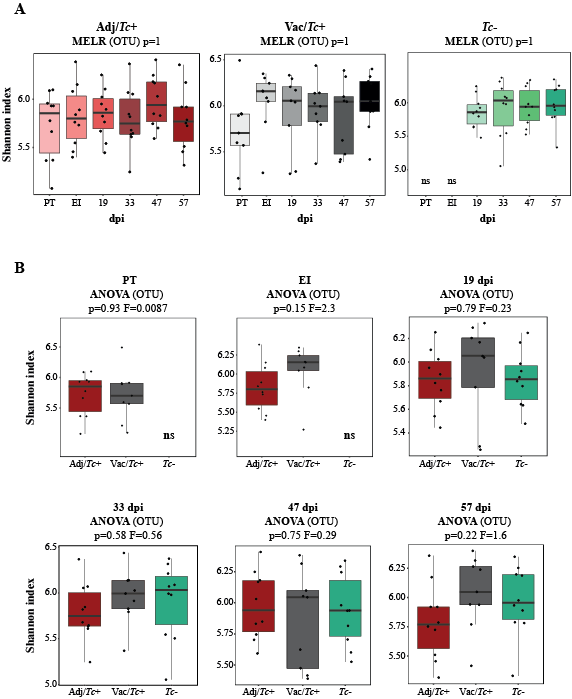
No statistically significant differences in gut microbial alpha diversity were recorded over time between vaccinated/ and unvaccinated/challenged animals, nor between these and uninfected control sheep, in accordance with previous studies conducted in ruminants and other large herbivores colonised by GI helminths, e.g. goats infected with the abomasal strongyle Haemonchus contortus [42], partially immune cattle challenged with infective larvae of Ostertagia ostertagi (an abomasal nematode whose life cycle and biology closely reflect those of T. circumcincta in sheep) [41] and horses harbouring large-intestinal strongyles [46–47]. On the other hand, throughout the course of the study, the gut microbiota of sheep challenged with T. circumcincta (irrespective of prior immunisation) displayed a progressively decreasing beta diversity, that reached statistical significance when the microbial profiles of samples collected prior to challenge infection were compared to that of samples collected at the end of the study period. This finding may result from the rise in pH of the gastric environment that follows the establishment of T. circumcincta [48], which is likely to enable a greater survival of selected ruminal bacteria (e.g. obligate anaerobes) transiting through the abomasum of infected ruminants [48]. In the present study, obligate anaerobes that were significantly expanded in the gut microbiota of T. circumcincta infected animals, compared to uninfected animals, included members of the Prevotellaceae, and Porphyromonas and Sutterella. Amongst these, Prevotella spp. proliferate when the pH of the medium increases [49–51], and the expansion of this genus of bacteria was also reported in the abomasum of Haemonchus-infected goats alongside a significant pH increase [42]. Prevotella spp. are key metabolisers of peptides and carbohydrates [52–55]; thus, the expansion of this genus of bacteria hadbeen hypothesized to serve as a compensatory mechanism to counteract the protein loss caused by abomasal helminth infections [42]. Nevertheless, the significant increase of Prevotella observed in the faeces of parasitized animals, together with the simultaneous expansion of other bacterial taxa, e.g. Porphyromonas spp., is likely to contribute to the pathophysiology of Teladorsagia ovine infection. Notably, these two taxa are common and generally harmless members of the resident gut flora that, under particular environmental conditions, or in individuals carrying selected genetic mutations, may become pathobionts [56–57] and cause immune-mediated diseases [58–66]. Furthermore, the expansion of the genus Sutterella in Tc+ sheep could also contribute to abomasal inflammation upon infection [67], though the mechanisms through which this taxon could enhance local inflammation are still not fully understood [68–70].
Indeed, the molecular mechanisms underlying pathobiont-mediated pathology are diverse, and often involve positive feedback loops that exacerbate inflammation (reviewed by [57]). One known mechanism involves loss of tolerance to otherwise innocuous microbes, and subsequent activation of microbiota-specific pro-inflammatory T cells (of the Th17 and Th1 subsets) in the presence of a heterologous GI infection [71–72]. The mucosal immune responses activated upon T. circumcincta infection are complex, and involve the activation of mixed Th1/Th2/Th17 cell populations [73–74]; in particular, a study of the transcriptome of the abomasal epithelium in response to chronic T. circumcincta infection showed that polarisation towards the Th17 subset is determinant for susceptibility to parasite colonisation (leading to gross mucosal inflammation), whilst the local immune responses of ‘resistant’ animals are polarised towards a Th2-dominated phenotype [73–74]. In these animals, mucosal histopathology in response to infection is limited [73]. The immune-molecular interactions that determine Th cell polarisation towards the Th17 or Th2 phenotype are still unclear [75–76]. Similarly, whether Th17-skewed responses in susceptible animals are fully addressed to the invading parasites or also to the changing microbiota is currently unknown. Indeed, our data suggest that alterations of the abomasal environment induced by T. circumcincta, including increased pH, epithelial permeability and mucosal inflammation [18], could promote the expansion of selected populations of bacteria that, in a positive loop, could trigger the onset of collateral, pathobiont-dependent inflammation. However, given that changes in the gut microbiota are likely triggered by the increase of the abomasal pH that follows worm establishment [42,48], the mechanisms underlying differences in overall pathology between susceptible and resistant sheep (displaying Th17- and Th2-dominated immune responses, respectively) are still unclear (cf. [73–74]).
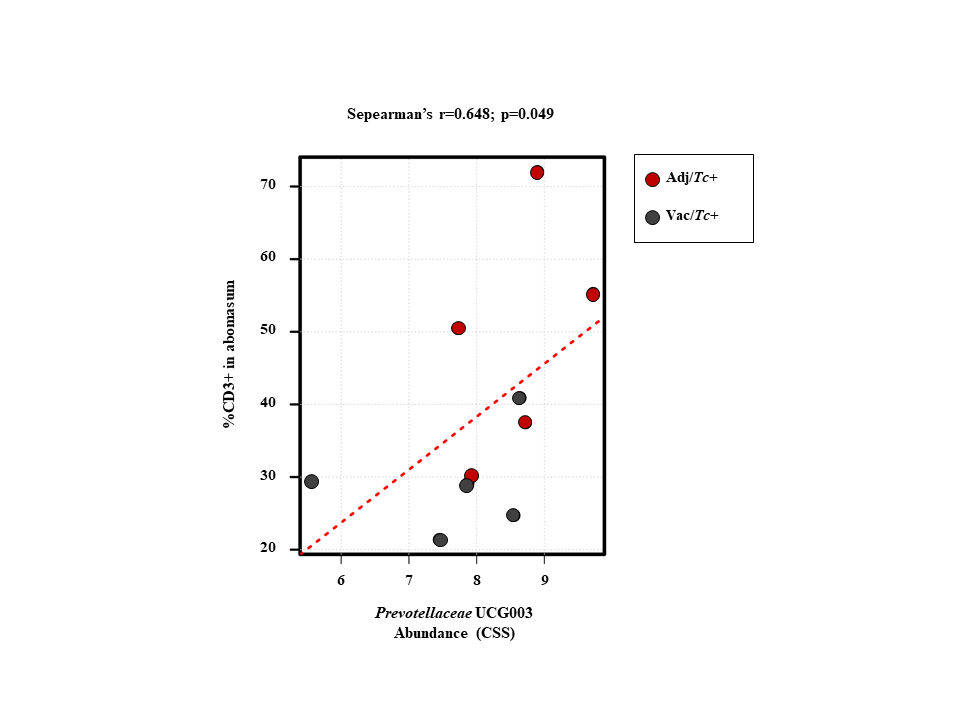
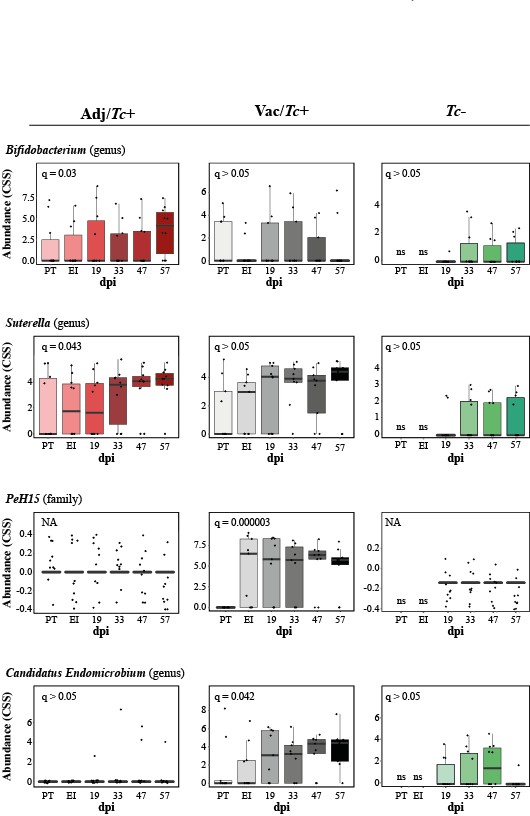
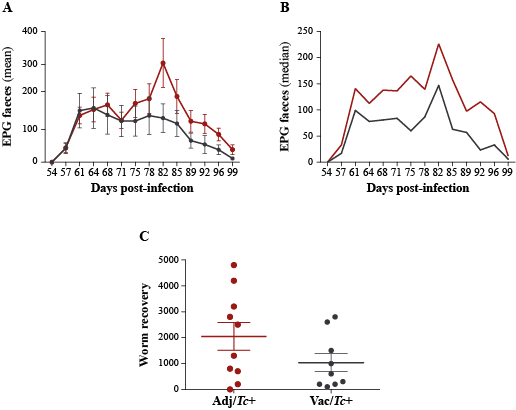
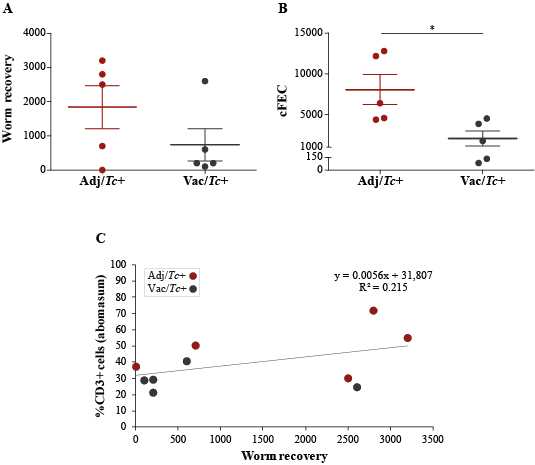
In the present study, two animal groups with different Teladorsagia worm burdens, i.e. the Adj/Tc+ and Vac/Tc+ animals, were investigated. Immunisation with a cocktail of recombinant parasite antigens resulted in a 56% reduction in mean peak faecal egg output and a 30% reduction in mean cumulative faecal egg output (cf. [23]). Both of these measurements are key indicators of the dynamics of the increase and decrease of parasite burden during the challenge period, whereas worm burden at post mortem is indicative of the number of worms that had survived till the end of the study. Worm burden measured at post mortem following a prolonged trickle infection protocol is therefore strongly influenced by factors such as the lifespan/rate of turnover of worms and interactions with the developing, exposure-induced, natural immunity [23]. In order to gain further insights into the immune microenvironment that accompanies parasite infections and the changing abomasal microbiota in naïve and immunised animals, local populations of mucosal T cells were studied via quantitative tissue microscopy, removing any reliance upon subjective observations and representative images. The results of our analyses indicate that the number of T lymphocytes infiltrating the abomasal tissue is directly proportional to worm burdens. Since infiltration of T cell populations in the abomasal mucosa has been associated with tissue damage linked to developing larvae [73], the enhanced recruitment of T cells into the abomasal mucosa of Adj/Tc+ sheep may result from higher loads of establishing parasites in this group compared with the Vac/Tc+ counterpart; therefore, under the experimental conditions of this study, assessing the roles that vaccination-induced local immunity and differences in worm burdens might play in protecting the host from microbiota-dependent pathology is challenging. Indeed, the gut microbial profiles of Adj/Tc+ and Vac/Tc+ sheep were largely similar and the effect of vaccination in the current trial was relatively small; thus, whilst significant expansion of the PeH15 family and of the Candidatus Endomicrobium genus of bacteria was detected in vaccinated animals, any speculation on the potential links between the abundance of these taxa and the immune features of the abomasal mucosa is currently unwarranted. However, a significant (weak) correlation was observed between numbers of T lymphocytes and the abundance of Prevotellaceae UCG003. This genus of bacteria was significantly expanded in both Adj/Tc+ and Vac/Tc+ sheep over the course of parasite infection; nevertheless, it was significantly more abundant in the gut microbiota of Adj/Tc+ than in that of Vac/Tc+ animals at the end of the study. The limited availability of sheep-specific antibodies for phenotypic characterisation of T lymphocytes via immunofluorescence labelling of formalin-fixed paraffin-embedded tissues impaired the identification of the specific T cell subsets whose abundance might correlate with cFEC and/or with that of selected gut microbial populations. Nonetheless, our data provide support to the hypothesis that, as for other GI helminth infections [71–72], microbiota-specific T cell responses might be activated following colonisation of the abomasum by T. circumcincta. Thus, future studies aimed to assess the contribution of each player, i.e. the macro- and microbiota, to the immunopathology of Teladorsagia infection are, in our opinion, warranted and timely.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}