First-round breeding using Chlamydomonas sp. KOR1 as the parent
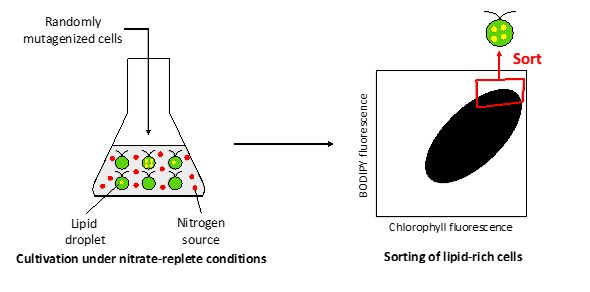
For one-step lipid production, microalgae that can accumulate lipids under nitrogen-replete conditions are needed. It was assumed that screening of lipid-rich cells from randomly mutated cells cultured in the presence of an abundant source of nitrogen would be useful to obtain such mutants. To examine this hypothesis, mutational breeding was performed using Chlamydomonas sp. KOR1 as the parent. KOR1 was irradiated with carbon ion (12C5+) beams to induce random mutations. The mutant cells were cultured under the nitrate-replete condition in MB 12N medium containing 2% (w/v) sea salt for 3 days. MB12N medium contains abundant NaNO3 (1588.4 mg/L) than previously used medium [11]. Nitrate concentration in the medium after 3 days cultivation was approximately 800 mg/L which shows nitrate in the medium was replete. Then, mutants were subjected to FACS-based screening of lipid-rich cells highly stained by BODIPY 505/515.
In the first sorting experiment, the chlorophyll fluorescence and BODIPY fluorescence intensity of most cells were 1.0 × 105 ~ 1.0 × 106 (a.u.) and 1.0 ×104 ~ 2.0 × 105 (a.u.), respectively (Fig. 1a). From these, 1,000 cells exhibiting high BODIPY fluorescence of 1.0 × 105 ~ 1.0 × 106 (a.u.) were sorted (Additional file1: Fig. S1 and Table S1). The sorted cells were subjected to repeated cultivation under nitrate-replete conditions and FACS-based sorting, for a total of five times. In the fifth sorting experiment, a cell population exhibiting higher BODIPY fluorescence than the original cells was observed (Fig. 1b), suggesting that mutant cells that accumulate lipids in the presence of nitrate were successfully screened. Then, the mutant colonies were isolated by inoculating the cells obtained in the fifth-sorting on Tris-acetate-phosphate (TAP) agar plates. For the secondary screening, 18 mutant strains were randomly selected (designated as KAC17s), cultured under nitrate-replete conditions for 3 days, at which time the lipid content was measured using gas chromatography-mass spectrometry (GC-MS). Among these mutants, KAC1710 exhibited the highest lipid content (18.3%), compared with lipid content of only 7.2% for KOR1 (Fig. 1c).
Second-round breeding using Chlamydomonas sp. KAC1710 as the parent
To further increase the lipid content under nitrate-replete conditions, second-round breeding was conducted using KAC1710 as the parent. Similar to the first-round breeding, KAC1710 was irradiated with a heavy-ion beam, and the nitrogen-conditioned screening of lipid-accumulating cells was performed using FACS. In the first sorting experiment, chlorophyll fluorescence and BODIPY fluorescence of most cells were 1.0 × 105 ~ 106 (a.u.) and 1.0 × 105 ~ 1.0 × 106 (a.u.), respectively (Fig. 2a). From these, 1,000 cells exhibiting higher BODIPY fluorescence of approximately 3.0 × 106 (a.u.) were sorted (Additional file1: Fig. S2 and Table S2). The sorted cells were repeatedly subjected to cultivation under nitrate-replete conditions and sorting of high BODIPY fluorescence cells, for a total of four times. In the fourth sorting experiment, most cells exhibited BODIPY fluorescence higher than 1.0 × 106 (a.u.) (Fig. 2b), suggesting that further improvement in lipid accumulation under nitrate-replete conditions was achieved. The cells obtained in the fourth sorting were then spread on TAP agar plates to isolate mutant strains. For the secondary screening, 15 mutant strains were randomly selected (designated as KAC18s), cultured under nitrate-replete conditions, and then their lipid content was analyzed by GC-MS. Among these mutants, KAC1801 exhibited the highest lipid content (23.1%), compared to 15.2% for the parental strain KAC1710 (Fig. 2c).
Increased formation of lipid droplets in KAC1710 and KAC1801
Transmission electron microscopy (TEM) analysis revealed that KAC1710 and KAC1801 cells accumulated lipid droplets even in the presence of nitrate, and KAC1801 accumulated more lipid droplets than KAC1710 (Fig. 3). In addition, KAC1710 and KAC1801 cells were larger in size than KOR1 cells, which might have been caused by the increased lipid droplet formation in these mutants. These results suggested that formation of lipid droplets under nitrate-replete conditions was increased by the two random mutagenesis and nitrate-conditioned screening steps.
Evaluation of lipid productivity of KAC1710 and KAC1801
To evaluate lipid productivity under nitrate-replete conditions, KOR1, KAC1710, and KAC1801 were cultured in MB 12N medium containing 2% (w/v) sea salt, and cell density, biomass, nitrate concentration in the medium, and lipid content were measured. Maximum cell density of KOR1, KAC1710, and KAC1801 during 6 days of cultivation was 8.0 × 106 cells/mL, 5.7 × 106 cells/mL, and 5.3 × 106 cells/mL, respectively (Fig. 4a). Maximum biomass of KOR1, KAC1710, and KAC1801 was 4.1 g-DCW/L, 3.6 g-DCW/L, and 3.3 g-DCW/L, respectively (Fig. 4b). These results suggest that delayed cell division in KAC1710 and KAC1801 led to the lower biomass production. In addition, nitrate consumption was decreased in KAC1710 and KAC1801 (Fig. 4c). KAC1710 and KAC1801 consumed 79.8% and 62.9% of the nitrate in the MB 12N medium, respectively, whereas KOR1 completely consumed the nitrate in the medium over 6 days. The lipid content of KAC1710 and KAC1801 at day 5 was approximately 19.1% and 26.6%, respectively, which was 1.5- and 2.1-fold higher than that of KOR1 (12.5%) (Fig. 4d). Although biomass production was reduced, lipid production and productivity of KAC1710 and KAC1801 were improved based on the increase in lipid content. Lipid production of KOR1, KAC1710, and KAC1801 at day 5 was 457.8 mg/L, 605.4 mg/L, and 810.4 mg/L, respectively (Fig. 4e). Lipid productivity of KAC1710 and KAC1801 at day 5 was 1.3-fold (121.1 mg/L/day) and 1.8-fold (162.1 mg/L/day) higher, respectively, compared to KOR1 (91.6 mg/L/day) (Fig. 4f). Thus, by increasing lipid accumulation, lipid productivity under nitrate-replete conditions was improved.
Lipid productivity of KAC1710 and KAC1801 under nitrate-deficient conditions was also investigated using MB 6N medium containing 2% (w/v) sea salt, which contains half the amount of nitrate compared to MB 12N medium (Additional file1: Fig. S3). Biomass production of KAC1710 and KAC1801 after 10 days of cultivation was 5.1 g-DCW/L and 4.4 g-DCW/L, respectively, whereas that of KOR1 was 5.7 g-DCW/L (Additional file1: Fig. S3a). Nitrate in the medium was completely consumed in 3 days by KOR1, whereas 4 days were required by KAC1710 and KAC1801 (Additional file1: Fig. S3b). The lipid content of KOR1, KAC1710, and KAC1801 at day 10 was 43.5%, 50.7%, and 37.1%, respectively (Additional file1: Fig. S3c); thus, under nitrate-depleted conditions, improvement was observed only in KAC1710. Also, lipid production of KOR1 and KAC1710 was 249.7 mg/L/day and 259.8 mg/L/day, respectively, whereas in KAC1801, it decreased to 163.5 mg/L/day (Additional file1: Fig. S3d).
{kind=link}