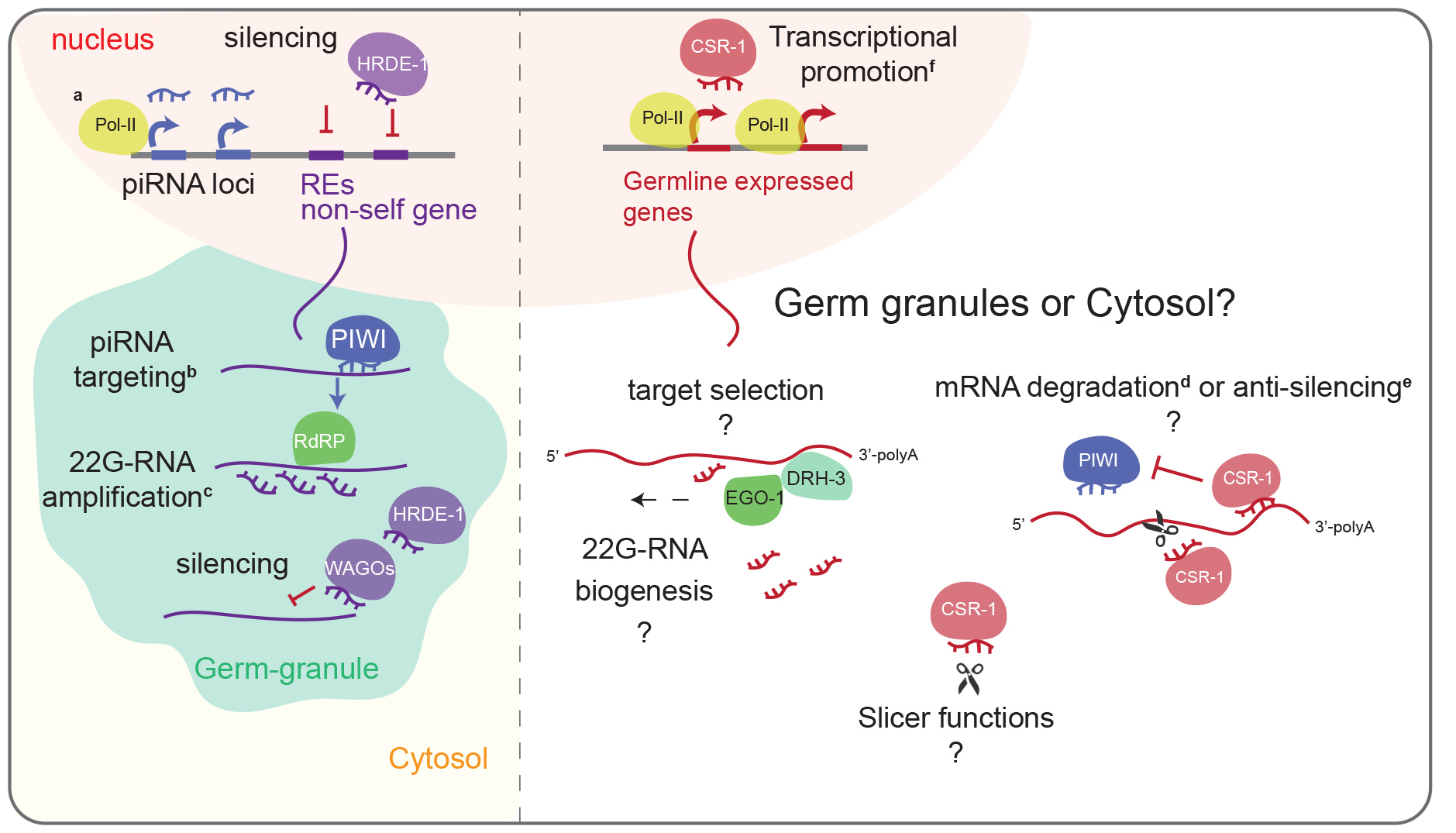
Defects in CSR-1 catalytic activity mainly impact 22G-RNA abundance
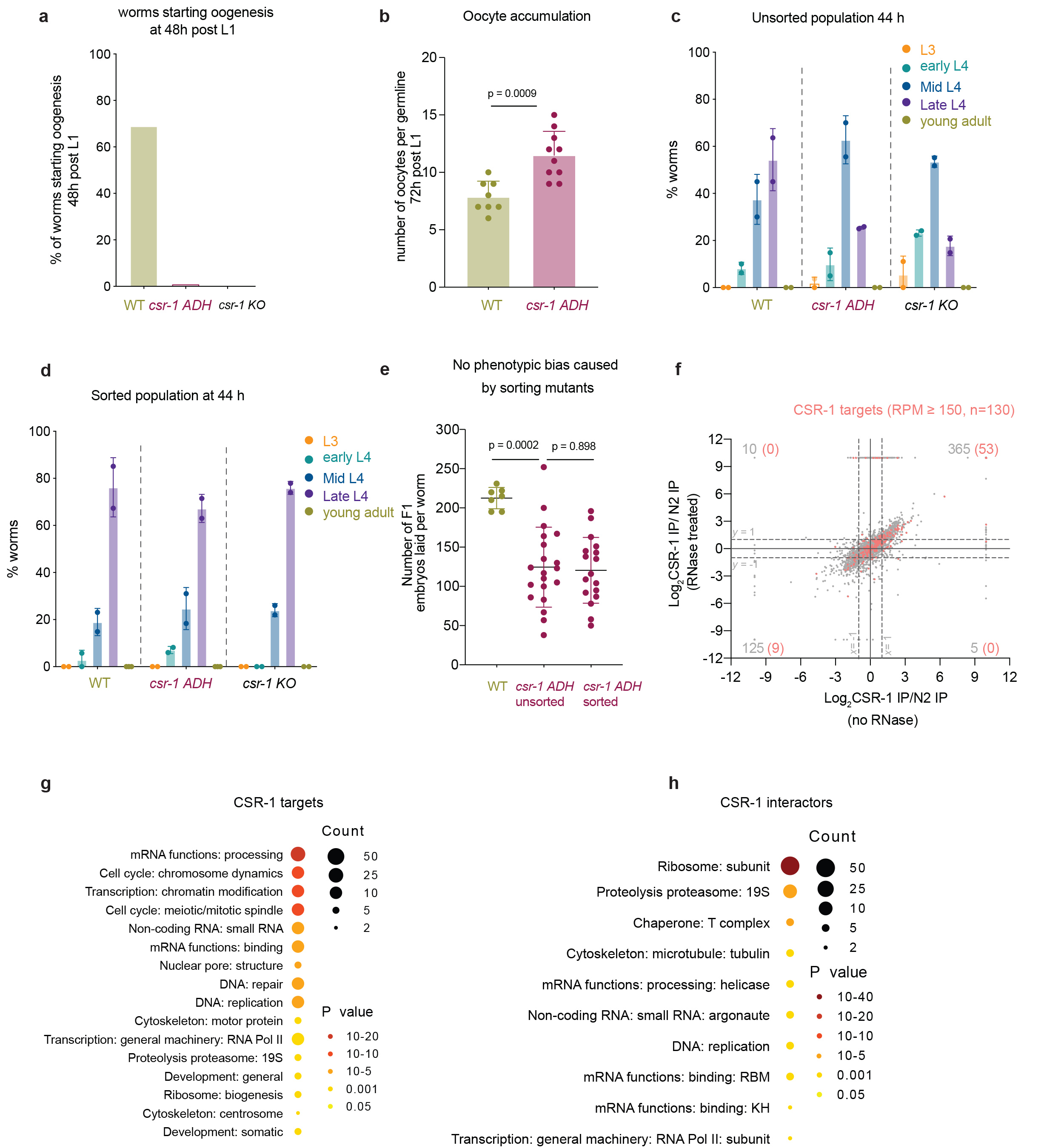
Worms lacking CSR-1 show downregulation13,20,21 whereas those expressing CSR-1 protein in which the catalytic DDH motif was mutated to ADH show upregulation of some targets22. The global impact of CSR-1 mutations on gene expression might depend on the developmental context and might be biased by developmental defects13,28. Indeed, we observed differences during oogenesis in the adult csr-1 catalytic mutant (csr-1 ADH) and knockout (csr-1 KO) worms marked by a delayed onset of oocyte production and increased accumulation of oocytes in germline at more advanced stages compared to wild-type (WT) (Extended Data Fig. 2a-c). To overcome this limitation, we developed a sorting strategy to obtain a synchronized population of WT and first-generation homozygotes for csr-1 KO or CSR-1 ADH strains using COPAS biosorter. Using this strategy, we enriched for larval stage late L4 worms which is characterized by a closed vulva and absence of oocytes and lacking the germline developmental abnormality (Extended Data Fig. 2d-e).
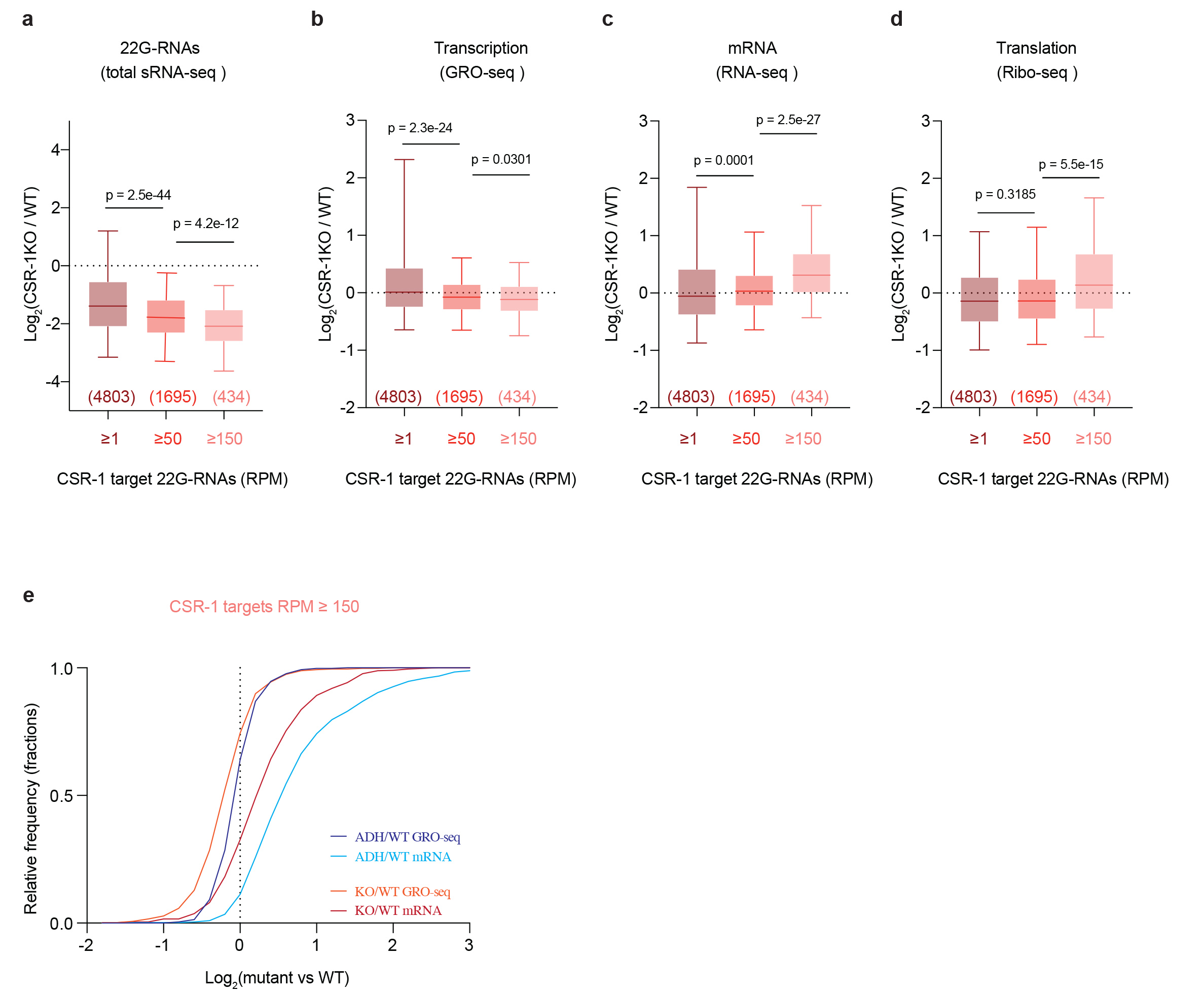
Next, to precisely evaluate the role of CSR-1, we measured small RNA accumulation (sRNA-seq), transcription (GRO-seq), mRNA stability (RNA-seq), and translation (Ribo-seq) in WT and mutant worms. In addition, to assess the direct effect of CSR-1 22G-RNAs on these processes, we sequenced the small RNAs bound to immunoprecipitated CSR-1 from similarly sorted late L4 worms to precisely identify the CSR-1 targets at the same developmental stage. We detected a total of 4803 genes with antisense 22G-RNAs loaded into CSR-1 (IP over input ≥ 2-fold enrichment and RPM ≥ 1 in each replicate of CSR-1 IP) (Supplemental Table 1). The CSR-1 catalytic mutant displayed a global loss of 22G-RNAs for the majority of CSR-1 targets (Fig. 1a, c). However, only 7.7% (n = 119) of CSR-1 targets with > 2-fold reduction of 22G-RNAs (n = 1536) showed increased mRNA levels (Fig. 1b), indicating that most mRNA targets are not destabilized by CSR slicer activity. The increase in mRNA and translational levels of the targets correlated with 22G-RNA levels in CSR-1 IPs in a dose-dependent manner (Fig. 1d, e) in agreement with a previous report22, but their transcription was unaffected (Fig. 1f). Therefore, we conclude that CSR-1 slices a subset of target mRNAs having abundant 22G-RNAs. Moreover, we found that the targets that are post-transcriptionally regulated by CSR-1 are enriched for mRNAs encoding CSR-1 interacting proteins, identified by mass spectrometry (MS/MS) (enrichment factor 6.1, p < 1.6e− 28) as direct, RNA-independent interactions (Extended Data Fig. 2g-i). Thus, CSR-1 slicer activity negatively regulates the expression of its own interactors, including CSR-1, suggesting a negative feedback loop.
Overall, these results suggest that the main role of CSR-1 catalytic activity is to control the accumulation of 22G-RNAs. In addition, CSR-1 post-transcriptionally regulates a small fraction of CSR-1 targets that have highly abundant 22G-RNAs.
CSR-1 protects a subset of oogenic enriched targets from piRNA-mediated transcriptional silencing
Similar to CSR-1 ADH worms, CSR-1 KO worms displayed a loss of 22G-RNAs as well as an upregulation of a subset of target mRNAs characterized by high abundance of 22G-RNAs (Extended Data Fig. 3a-d). However, the level of upregulation of CSR-1 target mRNAs was significantly lower in the csr-1 KO compared to the csr-1 ADH, possibly due to decreased transcription (Extended Data Fig. 3e). Indeed, we found that a subset of target genes displayed downregulated transcription and reduced mRNA levels in the KO compared to WT, but these were unaffected in the CSR-1 ADH (Fig. 1g). The majority of these genes (53%) were enriched for oogenic mRNAs (see Supplemental Table 1 for gene list). Given that CSR-1 is proposed to protect germline transcripts from piRNA-mediated silencing, we hypothesized that in the csr-1 KO, piRNAs can trigger the loading of 22G-RNAs into the nuclear Argonaute HRDE-1 resulting in the reduced transcription of this subset of CSR-1 targets. Indeed, HRDE-1 loads 22G-RNAs from transcriptionally downregulated CSR-1 targets in the csr-1 KO (Fig. 1h). Overall, these data suggest a non-catalytic role of the CSR-1 protein in protecting a subset of oogenic targets from piRNA-mediated HRDE-1 transcriptional silencing.
CSR-1 catalytic activity is required for biogenesis of 22G-RNAs on the coding sequence of target mRNAs
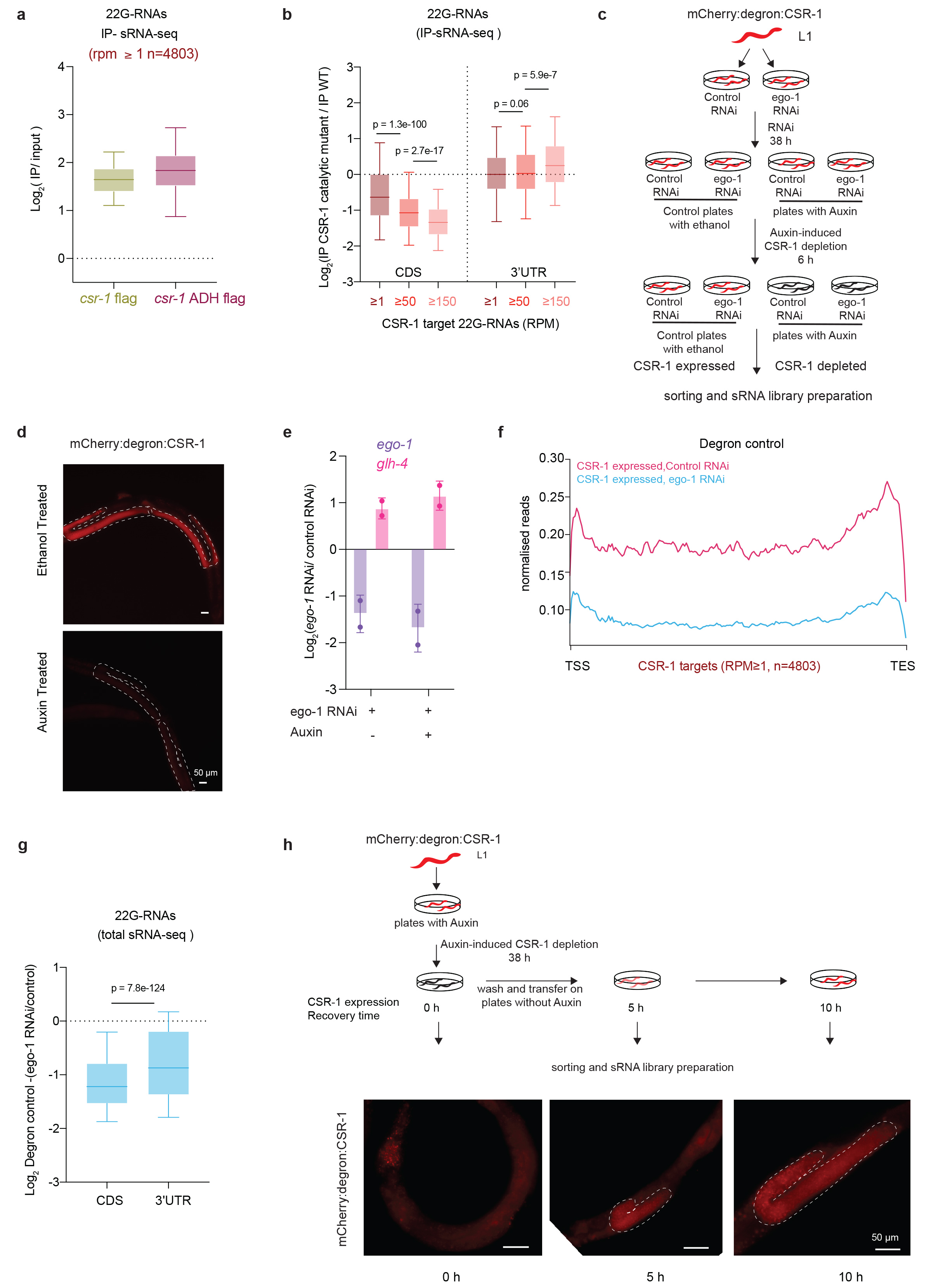
The global reduction of CSR-1-bound 22G-RNAs observed in CSR-1 mutants suggests that CSR-1 catalytic activity is required for 22G-RNA loading or biogenesis. Immunoprecipitation with WT or CSR-1 ADH proteins did not show any loss of binding of 22G-RNAs (Extended Data Fig. 4a), suggesting that catalytic inactive CSR-1 can still bind the 22G-RNAs produced in the mutant. We then investigated the distribution of CSR-1-bound 22G-RNAs along the target gene bodies. We found that 22G-RNAs are synthesized only at 3' untranslated region (3'UTR) of their target mRNAs in the csr-1 catalytic mutant, indicating that the RdRP fails to synthesize 22G-RNAs on the coding sequence (CDS) (Fig. 2a-c). The CSR-1 catalytic mutation does not impair the loading of these 22G-RNAs generated from 3'UTR (Fig. 2a, c and Extended Data Fig. 4b). However, it fails to produce and load small RNAs from CDS (Fig. 2a-c and Extended Data Fig. 4b).
The RdRP EGO-1 has been proposed to exclusively synthesize CSR-1-bound 22G-RNAs12,13,23. To understand whether the small RNAs produced on the 3’UTR in the absence of CSR-1 protein or its catalytic activity are synthesized by EGO-1, we efficiently depleted CSR-1 using an auxin-induced degradation system, combined with ego-1 RNAi knockdown (Extended Data Fig. 4c-e). First, we confirmed that CSR-1 22G-RNAs were depleted on CDS and enriched on 3'UTR upon auxin-induced CSR-1 depletion (Fig. 2d, e), suggesting that the effect on CSR-1 22G-RNAs was not caused by a gain-of-function mutation in CSR-1 ADH. Next, we observed decreased 22G-RNAs from both CDS as well as 3'UTR upon ego-1 RNAi knockdown (Fig. 2d, f, and Extended Data Fig. 4f, g), implying that EGO-1 is exclusively responsible for the synthesis of the CSR-1 22G-RNAs in both WT and the csr-1 mutants. However, the catalytic activity of CSR-1 is required to efficiently generate EGO-1-dependent 22G-RNAs along the coding sequences of target mRNAs.
Finally, we tested whether the restored expression of CSR-1 is sufficient to generate EGO-1-dependent 22G-RNAs on the gene body. For this purpose, we depleted CSR-1 by auxin-induced degradation for 38 h after hatching (0 h recovery) and then reintroduced CSR-1 by recovering expression for 5 and 10 hours (Extended Data Fig. 4h). As expected, the depletion of CSR-1 caused a loss of 22G-RNA accumulation on the CDS (Fig. 2g and Extended Data Fig. 4h - see 0 h recovery). However, upon reintroduction of CSR-1 expression (5 and 10 h recovery), we observed a steady increase of 22G-RNAs, mainly on the CDS (Fig. 2g, h). The lack of complete recovery of 22G-RNAs could be due to the accumulation of germline defects as a result of CSR-1 depletion during the initial period of germline development.
Overall, these data demonstrate that EGO-1 can be recruited on the 3'UTR of target mRNAs and initiate the production of 22G-RNAs. However, CSR-1-mediated slicing of mRNAs is required to template the production of small RNAs on the gene body.
Biogenesis of CSR-1 22G-RNAs and the regulation of their targets occurs in the cytosol
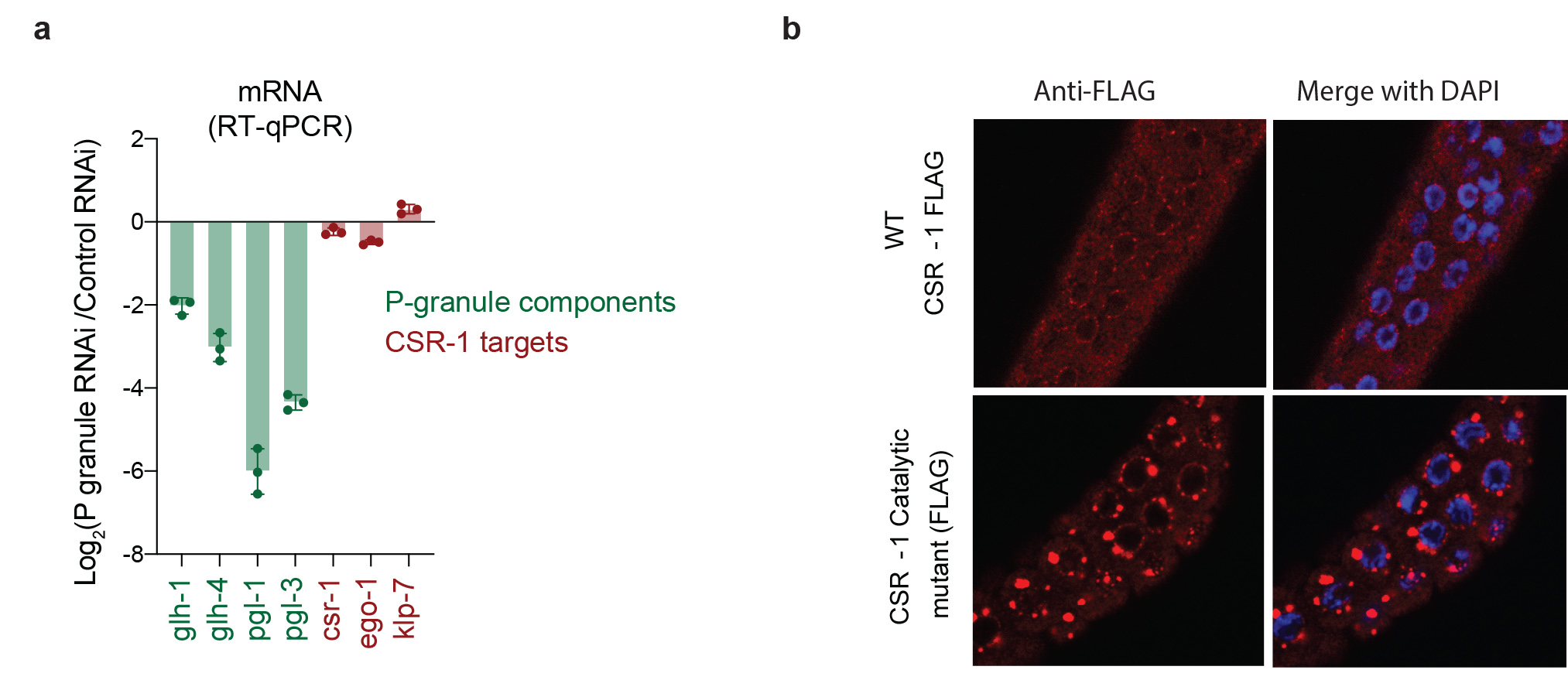
PIWI and RNAi biogenesis factors are known to localize in perinuclear condensates, called germ granules, and these germ granules have been proposed to be the site for biogenesis of 22G-RNAs24,29−31. CSR-1 and EGO-1 localize in both cytosol and the germ granules13, suggesting that the biogenesis of CSR-1 22G-RNAs might also occur in these organelles. To test this possibility, we used RNAi to simultaneously deplete four core components of germ granules (pgl-1, pgl-3, glh-1, and glh-4)32, called P granules (Extended Data Fig. 5a). This treatment was sufficient to disrupt the localization of the GLH-1, PIWI and CSR-1 in germ granules (Fig. 3a). However, the cytosolic localization of CSR-1 remained unaffected (Fig. 3a).
Next, we evaluated the effects of loss of germ granule localization of PIWI and CSR-1 on 22G-RNA biogenesis. As expected, piRNA-dependent 22G-RNAs were globally depleted upon P granule RNAi treatment (Fig. 3c). Surprisingly, CSR-1 22G-RNAs were unaffected upon P granule RNAi treatment, despite the loss of perinuclear CSR-1 germ granule localization (Fig. 3b, c). Furthermore, CSR-1 targets were not upregulated upon P granule RNAi (RNA-seq data from33 (Fig. 3d and Extended Data Fig. 5a). These results highlight that CSR-1 22G-RNA biogenesis occurs in the cytosol independently of germ granules.
Translating mRNAs serve as the template for 22G-RNA biogenesis
Our data so far suggest that CSR-1 22G-RNAs are generated in the cytosol. Consistent with CSR-1 localization in the cytosol and germ granules, we identified ribosomal and ribosomal-associated proteins, which are enriched in the cytosol, and germ granule components in our IP-MS/MS as direct CSR-1 interactors (Fig. 4a and Barucci et.al34). Moreover, CSR-1 ADH showed reduced co-purification of ribosomal proteins and increased co-purification of germ granule components, compared to CSR-1 WT (Fig. 4b) and localizes primarily in germ granules (Extended Data Fig. 5b).
Based on these data, we hypothesized that 22G-RNAs are synthesized in the cytosol, using translating mRNAs as templates. To test this hypothesis, we mapped the distance between the start of the 29-nucleotide Ribosomal Protected Fragments (RPF)35 and the 5' end of CSR-1 22G-RNAs (Fig. 4c). We observed periodicity of 3 nucleotides in phase with ribosomes (Fig. 4d), indicating that the synthesis of CSR-1 22G-RNAs occurs on mRNA templates engaged in translation. In contrast, the HRDE-1 loaded 22G-RNAs of P granule dependent piRNA targets (Supplemental Table 1) did not show phasing with ribosomes as observed due to a lack of 3 nucleotide periodicity (Fig. 4c and d), in agreement with the fact that P granules are devoid of translating mRNAs36,37.
Altogether these results suggest that CSR-1 cleaves actively translating mRNAs, which become the template for EGO-1-mediated synthesis of 22G-RNAs on the coding sequence of mRNA targets.
mRNA translation antagonizes CSR-1 22G-RNA biogenesis
EGO-1 mediated synthesis of CSR-1 22G-RNAs does not occur on every germline mRNA at similar levels, and we found that the levels of 22G-RNA are independent of the levels of the mRNA template (Extended Data Fig. 6a). Given our observations that actively translating mRNAs serve as the template for CSR-1 22G-RNAs, we hypothesized that the translation efficiency (TE) of germline mRNAs impacts CSR-1 22G-RNA biogenesis. To test this hypothesis, we calculated the TE of CSR-1 targets using the Ribo-seq and RNA-seq data from WT worms at the late L4 stage. We observed that levels of CSR-1 associated 22G-RNAs produced from a target mRNA were inversely correlated with their TE (Fig. 5a), suggesting that translation antagonize the biogenesis of CSR-1 22G-RNAs.
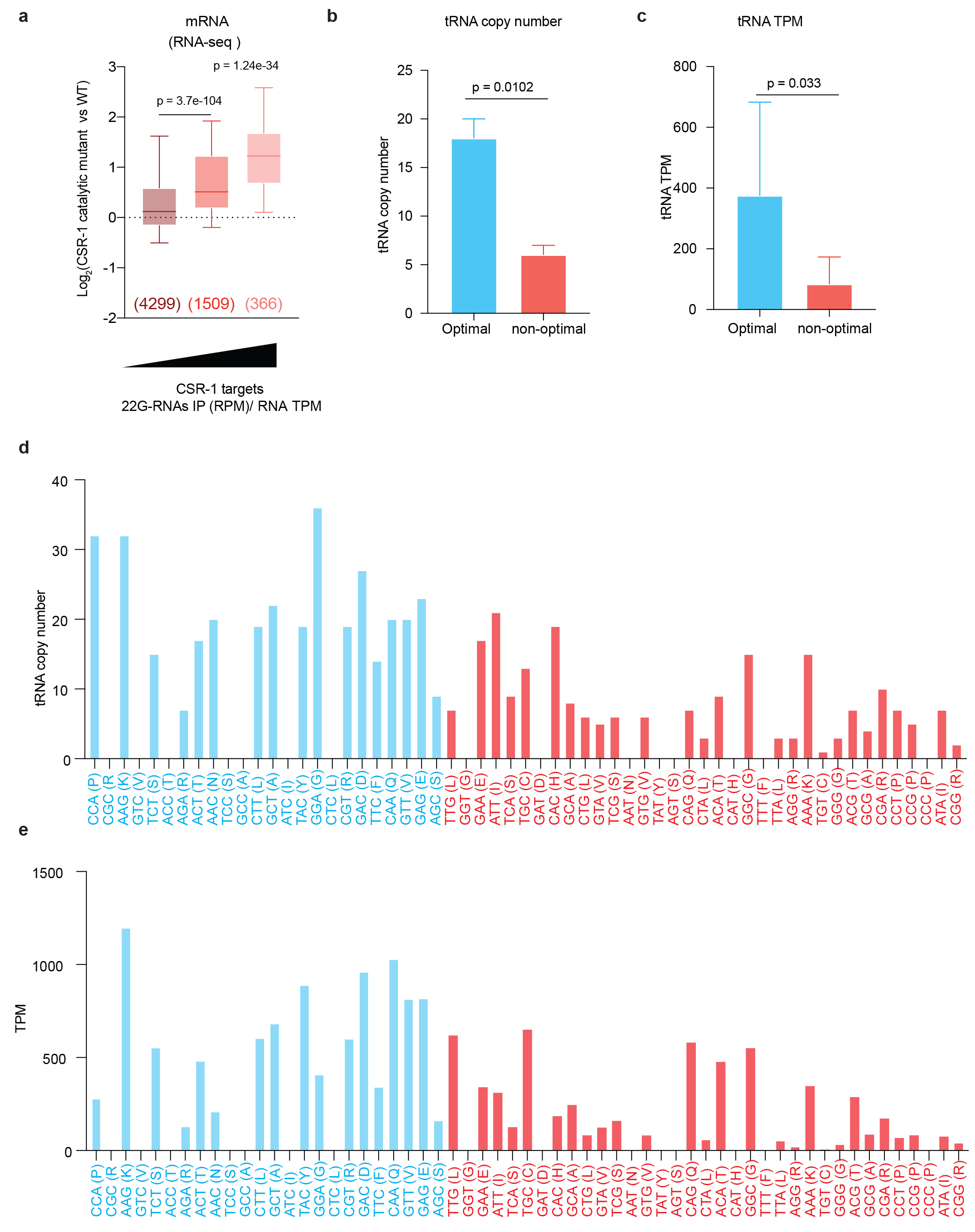
Codon usage and the availability of the tRNA pool influence TE38,39. Therefore, we investigated whether these mechanisms affect the biogenesis of CSR-1 22G-RNAs. We determined optimal and non-optimal codons using our experimental data from Late L4 staged worms. First, we calculated the normalized average relative synonymous codon usage (RSCU) for genes for different categories of high or low TE (Fig. 5b). Codons showing enrichment in genes with high TE (log2TE ≥ 3) were considered optimal codons, and the ones under-represented were considered non-optimal codons (Fig. 5b). We confirmed that our classification of optimal/non-optimal codons correlated with tRNA copy number (Fig. 5d, Extended Data Fig. 6b, d) and tRNA pool available in the late L4 worm population (44 h) as measured by GROseq (Fig. 5e, Extended Data Fig. 6c, e). We noticed that for codons with no tRNA cognates and requiring tRNA binding by wobble pairing, all optimal codons end with C, and non-optimal with U. Translation elongation is lower for those ending with a U40.
We then evaluated the codon usage of CSR1 targets by comparing their normalized average RSCU to highly translated mRNAs. We found that non-optimal codons were enriched and optimal codons were depleted in CSR-1 targets, suggesting that this might be an encoded feature of mRNA targets influencing the priming of 22G-RNA synthesis (Fig. 5c). Non-optimal codons are known to promote ribosome stalling41–43. To map differences in 22G-RNA biogenesis on sequences with optimal or non-optimal codons, we divided RPFs into two categories based on the presence of either an optimal or non-optimal codon at the A and P sites of the ribosome and then mapped the distance between 5’ of 22G-RNAs and RPFs. We observed a peak for the 5' end of 22G-RNAs downstream of RPF (29th position) when the A and P sites of the ribosomes are occupied by a non-optimal codon contrary to when optimal codons are present on A and P sites which show no bias (Fig. 5f). This result suggests that the 22G-RNA production is preferentially initiated downstream of ribosomes especially occupying bad codons, by CSR-1 mediated slicing and recruitment of EGO-1.
Altogether, these observations suggest that translation and ribosome position dictate the production of CSR-1 22G-RNAs.
Increasing the translation efficiency of CSR-1 target impairs CSR-1 22G-RNA biogenesis and function.
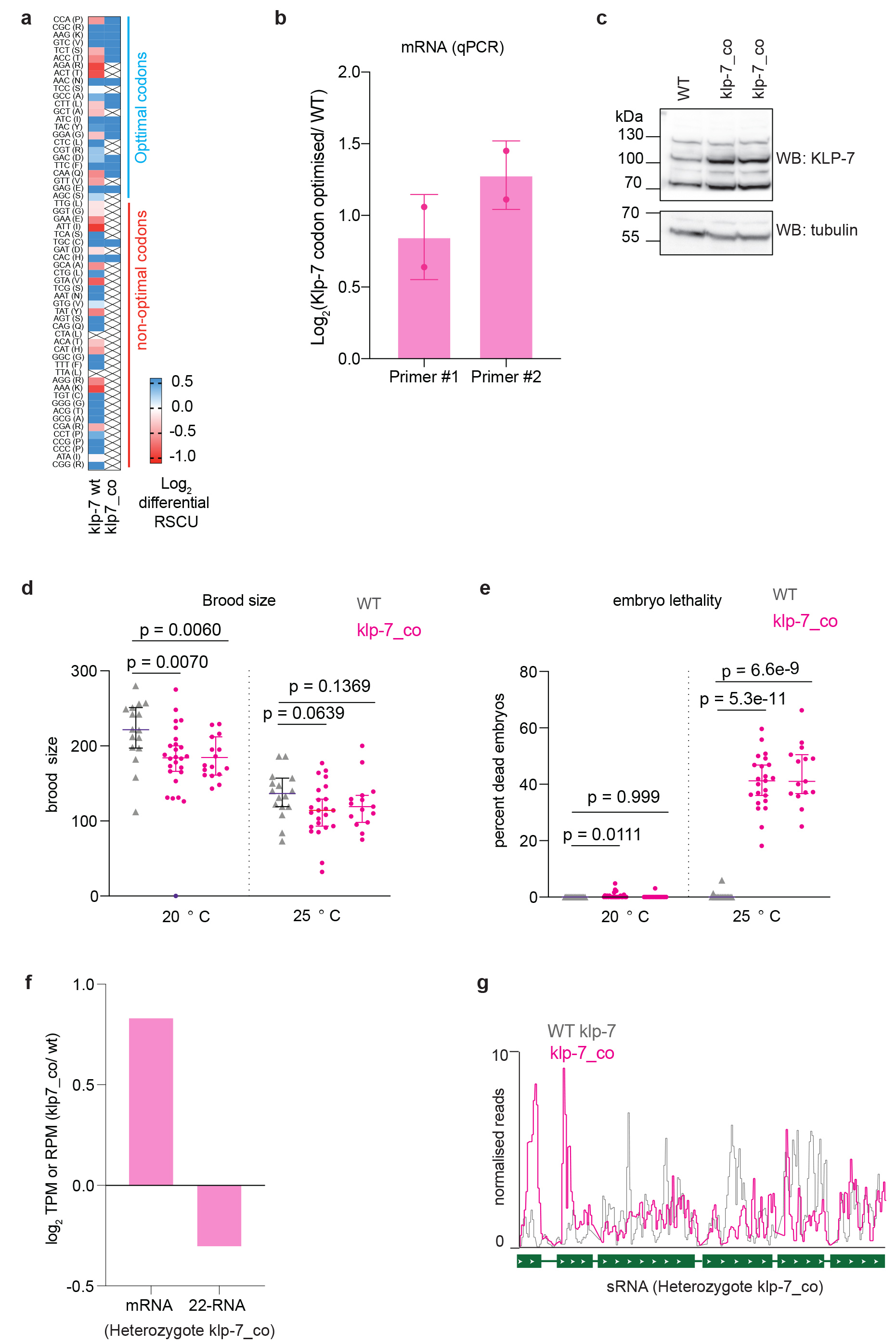
To determine whether non-optimal codons directly affect TE and CSR-1 22G-RNA biogenesis, we altered the coding potential of a CSR-1 target. We examined klp-7, which has the second-highest abundance of 22G-RNAs loaded by CSR-1 and is post-transcriptionally regulated by CSR-1. KLP-7 is a kinesin-13 microtubule depolymerase and is required for spindle organization and chromosome segregation44. Overexpression of KLP-7 in the csr-1 mutant has been shown to cause microtubule assembly defects22. klp-7 showed enrichment of non-optimal codons and depletion of optimal codons similarly to other CSR-1 targets (Extended Data Fig. 7a). We optimized the codon usage in klp-7 by incorporating exclusively synonymous optimal codons (Extended Data Fig. 7a). We used CRISPR-Cas9 to replace endogenous klp-7 isoform b with the modified klp-7 codon-optimized (klp-7_co) to avoid disrupting potential UTR-mediated regulation.
To ascertain whether codon optimization of klp-7 affected the TE, we performed RNA-seq and Ribo-seq from synchronized and sorted late L4 population (44 h). Indeed, we detected a 2-fold increase in the TE of klp-7 mRNA in the klp-7_co strain compared to WT (Fig. 6a). The TE of other CSR-1 targets remained unaffected in klp-7_co strain, indicating that the effects observed are specific to klp-7 mRNA (Fig. 6a). In addition, KLP-7 protein levels were increased in two independent lines of klp-7_co compared to WT, consistent with increased translation (Extended Data Fig. 7c). We then measured the level of 22G-RNAs antisense to klp-7 mRNA in the klp-7_co strain compared to WT, and we observed a 1.4-fold decrease in 22G-RNAs (Fig. 6a). The levels of 22G-RNAs for other CSR-1 targets remained unaffected (Fig. 6a). Further, the significant decrease in 22G-RNAs on the optimized klp-7_co allele was observed in exons 3–6 and was accompanied by an increase in ribo-seq peak height at those positions (Fig. 6b). The klp-7_co strain also showed increased klp-7 mRNA level compared to WT (Fig. 6a), and we confirmed this result by RT-qPCR (Extended Data Fig. 7b). These results suggest that CSR-1 targeting and regulation is impaired on klp-7_co mRNA. To validate this, we performed csr-1 RNAi and showed increased klp-7 mRNA levels in the WT strain but not in the klp-7_co strain (Fig. 6c), suggesting that CSR-1 activity is reduced at klp-7_co mRNA. The increased levels of klp-7 mRNA correlated with a reduction in brood size (Extended Data Fig. 7d) and higher embryonic lethality at 25 °C in klp-7_co strain compared to WT (Extended Data Fig. 7e), indicating the physiological relevance of klp-7 mRNA targeting by CSR-1. Finally, to rule out any difference in the production of either 22G-RNAs or mRNA levels due to possible developmental defects between klp-7_co and WT strain, we generated a heterozygote strain of klp-7_co with a fluorescent GFP marker on the balancer chromosome. We sorted heterozygote GFP positive worms with one copy of modified klp-7_co and one copy of WT klp-7 each and performed RNA-seq and sRNA-sEq. We observed similar results with a 1.8- fold increase in mRNA levels for klp-7_co compared to the WT klp-7 copy and a 1.25-fold decrease in 22G-RNA levels (Extended Data Fig. 7f-g). These results demonstrate that CSR-1 22G-RNA biogenesis and activity is reduced on mRNA templates with optimized codons and increased translation.
Altogether, these results suggest efficiently translating ribosomes block the access of CSR-1 to the mRNA template and thereby hamper 22G-RNA production and, in turn, affects gene regulation by CSR-1 during germline development.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}