NAD(P)H fluorescence lifetime imaging of murine intestinal environment
Using NAD(P)H-FLIM, we analyzed the general metabolic activity and specific enzymatic activity of the nematode H. polygyrus and of its natural environment, the luminal side of duodenum in mice. In order to access the intestinal niche of adult H. polygyrus nematodes, duodenum of infected C57/Bl6 mice was explanted and cut open to expose its luminal side. Here, the parasites, coiled around intestinal villi, persist over several weeks (Fig. 1a).
The fluorescence of the ubiquitous coenzymes NAD(P)H was detected at 466±30 nm after two-photon excitation at 760 nm. The coenzymes present both in nematodes and in enterocytes and cells of the lamina propria of murine intestinal villi, were also used to visualize worm and host tissue architecture. Based on NAD(P)H fluorescence images, we used a UNet-based pixel classification algorithm to segment the nematodes as well as epithelial layer and lamina propria of the murine intestinal villi and to exclude background regions (Fig. 1b; Material and Methods).
The time-resolved NAD(P)H fluorescence decay acquired in each pixel of the image (Fig. 1c) was recorded by time-correlated single-photon-counting34,35 and evaluated using the phasor approach to FLIM36, generating the real and imaginary phase maps and the average fluorescence lifetime (t) map (Fig. 1c).
The time-resolved NAD(P)H fluorescence signal provided further information on the molecular environment in cells and tissue. The coenzymes NADH and NADPH are able to bind to diverse enzyme and participate in this way to different metabolic pathways and to reductive biosynthesis within cells29,37-40. The fluorescence lifetimes of both unbound NADH and unbound NADPH lay at ≈450 ps41,42, both being the average of the fluorescence lifetime over two folding states of each type of coenzyme molecule (≈200 ps and ≈700 ps)43. When bound to enzymes, the fluorescence lifetime of NAD(P)H becomes longer (≈2000 ps), and it strongly depends on its enzymatic binding site. Taking into account the abundance of NAD(P)H-dependent enzymes in living cells (mostly valid across species), we previously developed a systematic frame work to evaluate NAD(P)H-FLIM data44. This approach allows us to assess the general metabolic activity, i.e. the percentage of bound NAD(P)H from the total NAD(P)H - activity map and the preferential specific enzymatic activity - enzyme map - in each pixel of the acquired image, both for the worm and for the host tissue (Fig. 1d). High values in the activity map indicate high metabolic activity, whereas low values indicate decreased metabolism, possibly associated with dormancy, low temperature or cell death45. The preferential binding of NAD(P)H to specific enzymes in the enzyme map indicates various metabolic pathways within cells and tissues, e.g. a preferential activation of lactate dehydrogenase (LDH) is associated with anaerobic glycolysis. The activation of pyruvate dehydrogenase (PDH) is associated with oxidative phosphorylation, while the activation of NADPH oxidases (NOX/DUOX enzymes) indicates metabolic defense, i.e. via oxidative burst. Thus, the heterogeneity of dominant metabolic pathways in worm and host, with high axial (z) resolution is monitored (Fig. 1e).
NAD(P)H metabolism of explanted duodenum
By performing NAD(P)H-FLIM in healthy, explanted duodenum (Fig. 2a) of C57/Bl6 mice, in RPMI medium, at 37°C, we assessed the metabolic activity of intestinal epithelial cells (EC) and of cells within the lamina propria (LP), respectively, as segmented from 3D NAD(P)H fluorescence images using the previously described pixel classification algorithm (Fig. 2b,c).
During imaging over up to 3 hours after explanting the duodenum, the general metabolic activity of the EC was 78.4±5.5 % (n = 6 mice, field-of-view 500x500 µm³ over 50 to 110 µm depth). In some mice, the cells in lamina propria show a slightly lower metabolic activity as compared to EC, in most mice the activity levels were comparable for lamina propria and epithelium, resulting into an activity value of 76.3±5.2 % in LP (p = 0.5139). The variability of metabolic activity was slightly higher in lamina propria than in the epithelium, i.e. standard deviation of activity levels per mouse were 8.1 % in LP as compared to 6.3 % in EC. These results resemble our findings regarding metabolic activity in small intestine villi of anesthetized CX3CR1:GFP mice acquired using in vivo NAD(P)H-FLIM12.
The enzymatic activity profile in the explanted healthy duodenum shows a preference towards oxidative phosphorylation in epithelium, with local increased activation of NADPH oxidases (Fig. 2d,f). As the first line of cellular defense in the intestine, EC show a high expression of NOX4 and NOX212. Both NOX2 and NOX4 activation leads to oxidative burst, i.e. massive extracellular reactive oxygen species (ROS) generation needed in the defense against pathogens. In addition, NOX4 activation is used for intercellular communication among intestinal epithelial cells46, in support of host defense. In contrast, the cells in LP balance anaerobic glycolysis and oxidative phosphorylation for energy production, with a slight preference towards anaerobic glycolysis. The LP cells show only few isolated spots of NADPH oxidase(s) activation, presumably NOX2 in phagocytes, significantly lower than the NOX activation observed in epithelium (Fig. 2e,f). These data are also in good agreement with the findings of analogous intravital experiments12.
Changes in metabolic activity in the intestinal villi during H. polygyrus infection
NAD(P)H fluorescence images of duodenum tissue from mice infected with H. polygyrus revealed a disorganization of the typical villi architecture in the vicinity of nematodes (Fig. 3a, Suppl. Fig. 1). Consequently, segmenting epithelium and lamina propria, as described for the healthy duodenum, was not always possible in the infected gut and we refer in all following data to intestinal tissue, without differentiating between epithelium and lamina propria.
When compared to healthy duodenum (77.3±5.2 %), NAD(P)H-FLIM analysis of infected duodenum tissue, 6 to 14 days post infection (acute infection), revealed a similarly high metabolic activity (74.0±7.6 %, n = 9 mice, Fig. 3c,d), with high heterogeneity within the villi (s.d. 9.6 %). A preference towards oxidative phosphorylation was observed in all mice (Fig. 3b,e,f). In contrast, at an earlier time point of infection, at day 6 after infection, where most parasites still reside in the submucosa, the typical villus morphology is preserved. At this time point during infection, in some areas in one mouse, a preference towards anaerobe glycolysis was observed (Fig. 3b, left image), however, in most duodenal villi, oxidative phosphorylation prevailed (n = 2 mice, Fig. 3e,f).
In line with the expected time course of immunomodulation caused by nematodes in the host 3,5,8,47-50, NADPH oxidases activation in the villi is rather low, i.e. comparable to healthy tissue, at day 6 after infection but increases dramatically at day 10 and 14 after infection, i.e. during the acute phase, in the vicinity of parasites (Fig. 3e,f).
NAD(P)H metabolic and enzymatic activity of live H. polygyrus in the gut
Adult H. polygyrus nematodes residing in the intestinal lumen display active motility of loose ends of their body, i.e. the ends not being coiled around villi (Fig. 4a, Suppl. Movie 1, 2). This active, undulatory motion through the intestinal mucus (labelled by the lipophilic dye BODIPY1) indicates the vitality of the worms and probably is key for their locomotion upwards in the small intestine and their survival in the duodenal luminal niche. In order to understand the energetics governing their motility behavior, we aimed to investigate NAD(P)H metabolism of nematodes by NAD(P)H-FLIM.
Using the same pixel classification approach as previously described, the nematode hydrostatic skeleton and its cellular, NAD(P)H-rich tissue were segmented from the NAD(P)H intensity images of H. polygyrus in the murine duodenum (Fig. 4b). The resulting mask was used to generate metabolic and enzyme 2D and 3D maps of the worms (Fig. 4c, d) and to calculate the relative preferential activation of the different metabolic pathways (Fig. 4e). Interestingly, several worms showed extensive NADPH oxidase activation, presumably DUOX2 activation, as the only member of the NADPH oxidases family expressed in H. polygyrus51. The activation of DUOX2 was particularly high in the digestive system and may indicate parasite defense against nematode environment.
Metabolic and specific enzymatic activity of H. polygyrus depends on the specific phase during infection
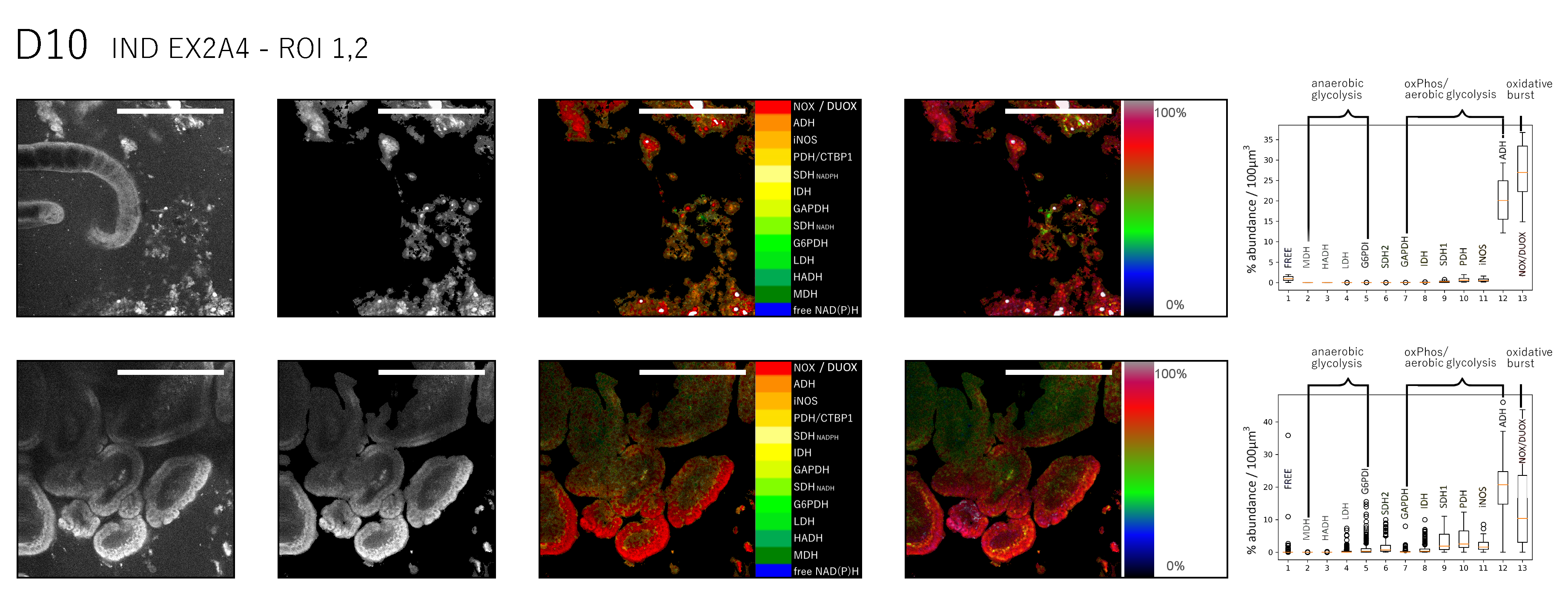
In order to understand if the translocation from the larval development in the submucosa to an adult worm in the gut lumen is associated with a specific metabolic pathway, we tracked the changes in NAD(P)H metabolism of H. polygyrus parasites between day 6 and day 14 after infection, using NAD(P)H-FLIM (Fig 5a, b, c).
As compared to the host intestinal tissue, nematodes show a lower metabolic activity (64.3±6.3 %, n = 16 worms from 14 mice, between day 6 to 14 after infection), with a similarly high heterogeneity per worm of 9.3 %. Interestingly, the general metabolic activity in worms slightly increases from day 6 to day 14 after infection, i.e. day 6 metabolic activity 58.6±2.2 % (2 worms), heterogeneity per single worm 6.1 %, day 10 61.1±9.9 % and 11.8 % (5 worms), day 12 64.5±5.8 % (4 worms) and day 14 73.1±5.9 % (5 worms) (Fig. 5d).
At day 6 after infection, H. polygyrus parasites are located in the intestinal submucosa. At this stage, they generate energy both by anaerobe glycolysis and by oxidative phosphorylation and show very low levels of oxidative burst (DUOX2 activation). 10 days after infection, the parasites start to emerge from the submucosa into the lumen. This process is associated with a metabolic shift towards anaerobe glycolysis, at still low DUOX2 activation. Between day 12 and day 14 after infection, the adult parasites shift their metabolism towards oxidative phosphorylation and show increased levels of DUOX2 activation (oxidative burst), indicating defense mechanisms, possibly against the host (Fig. 5e,f).
{kind=link}