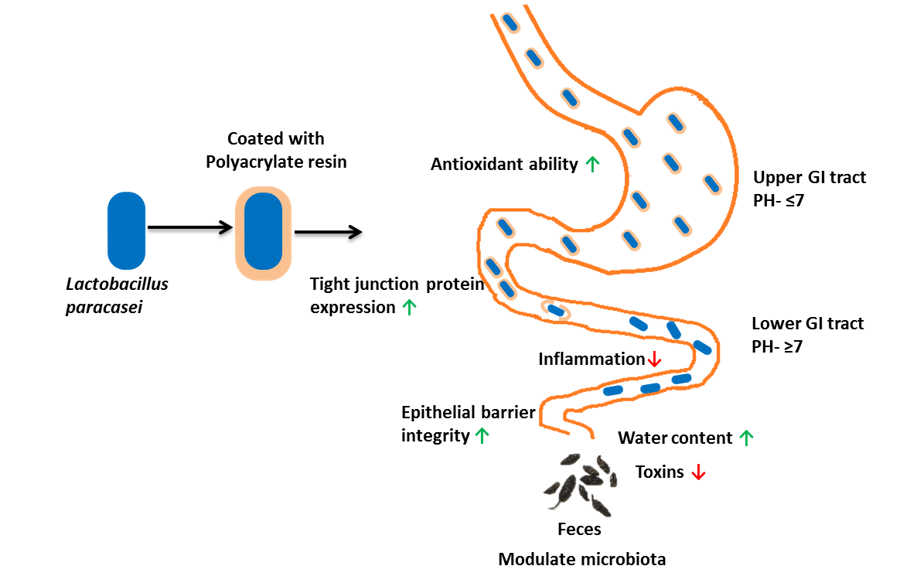
The purposes of microencapsulation are to mitigate the poor viability of probiotics due to the harsh environment on the upper part of the GI tract and to release probiotics at a controlled rate on the lower part for its beneficial action to the host. Previously, several microencapsulation methods and materials have been studied on different strains of probiotics to analyze their viability. Alginate-pectin microgel [34], chitosan-alginate [35], cellulose sulphate [36], etc., as encapsulating material protected different bacterial and fungal strains of probiotics from unfavorable gastrointestinal environment and improved the survival of bacterial cell, intestinal delivery and release resulting to several health benefits. However, pectin encapsulated probiotics did not enhance the effect of probiotics supplementation [26]. In our study, Polyacrylate resin was used as encapsulating material that can hold its structure at lower pH and release probiotic strain on the hindgut for the complete release of probiotic strain. Moreover, microcapsule formed improved overall gut performance.
Our encapsulated probiotics induced no comparable difference in body weight and food intake for four weeks. Similar results were recorded after the supplementation of different probiotic species [37, 38]. But, some strains of probiotics have shown notable improvement in growth performance [37, 39, 40].The moisture content in feces, which determines the softness or firmness, studies on probiotic species [41, 42]. The production of lactic acid [41] may be the reason for a higher moisture content that helps to alleviate constipation and induce bowel movement. Interestingly, we observed a significant increase in soleus weight that could be due to the variation in the gut microbiota and metabolites produced by them that influence the skeletal muscle mass [43]. But, for an accurate conclusion, further research is necessary.
The present study revealed that probiotic Lactobacillus species could improve intestinal morphology as we observed an increase in the villus height, decrease in crypt depth and V/C ratio on different intestinal tissues. They are the standard index for intestinal health and indicate enhancement in digestion and absorption by increasing the epithelium surface layer [44]. Similarly, intestinal epithelium layer represents the most important barrier against pathogenic molecules and bacteria. The integrity of the intestinal epithelial cell layer is maintained by adherens junctions, tight junctions, and desmosomes. Claudin, occludin and Zonula occludens (ZO-1) are majorly studied tight junction proteins [45]. The expression of claudin-1, occludin and ZO-1 were increased in the different intestinal tissue. Our result is consistent with other studies on a different strain of probiotics [46, 47].
LPS, also called endotoxins and TMAO, are intestinal microbiome-derived toxins correlated with inflammation, cardiovascular disease (CVD), and other diseases on the host [48]. The levels of these toxins were significantly reduced in the encapsulated group, suggesting Lactobacillus paracasei reduce the toxins production and the expression of markers of inflammation [49]. Short Chain Fatty Acids (SCFAs) are other key metabolites of microbiota in the colon. Lactate, short-chain hydroxyl-fatty acid, is produced by several bacterial species and converted to SCFA by lactate fermenting bacteria [50, 51]. The three monocarboxylic short-chain organic acids, lactate, acetate and propionate, can maintain immune and intestinal homeostasis by downregulating the pro-inflammatory response in intestinal epithelial cells, showing antimicrobial and anti-inflammatory effects [52, 53]. Our study showed a rise in the SCFA and lactate value in colonic, cecal and fecal content. Our study is consistent with several other probiotic species of Lactobacillus, Bifidobacterium, and Streptococcus, that produce Lactate and SCFA [54]. The modulation of microbiome metabolites is associated with a change in gut microbiota. Probiotics have a vital role in maintaining the gut microbiota by increasing the population of beneficial bacteria and reducing the pathogens by competing with them [55]. Previously, several studies were conducted about the impact of different strains of probiotics including Lactobacillus and it is reported that the probiotic modulates the microbiome on the different intestinal segments and control over microbes ecosystem [56]. In our study, we also found that the supplementation of encapsulated Lactobacillus paracasei changed the microbiome population.
The study on jejunal microbiota of chicken showed that probiotics could reduce the Shannon number on day 42 [39]. However, Zhang et al., 2015 investigated an increase in Shannon index in pro and pre encapsulated probiotics groups [20]. While another study on probiotics, Bacillus sp., increased Shannon index number in the colon but decreased in cecum against enterohemorrhagic Escherichia coli on mice [47]. Interestingly, we found a decrease in observed species and Shannon index. The treatment of encapsulated probiotic limited microbiome to predominantly Lactobacillus genus (0.4000 vs. 0.1952) and Lactobacillus murinus species (0.3194 vs. 0.1314), which may be the reason for the decrease in richness and diversity. We observed a clear difference between the control and encapsulated probiotic treated group, i.e., Beta diversity, which illustrates L. paracasei affects the microbiota in mice.
In our study, phylum Firmicutes was elevated and Bacteroidetes was slightly reduced in the treatment group, consistent with a previous study on probiotic species [39]. An increase in firmicutes and a decrease in Bacteroidetes phyla are correlated with increased absorption of nutrients [57], which indicated that supplementation of encapsulated Lactobacillus improves digestion by improving absorption. We also found upraised value in firmicutes to Bacteroidetes ratio (0.99 and 1.43) after supplementing the encapsulated probiotic, which is believed to be the marker for obese animals. However, we didn’t find any difference in weight gain in mice between those groups. A recent study has presented that this biomarker is still difficult to associate with the weight and health of an individual [6]. Lachnospiraceae family can be beneficial to the host as its members are chiefly responsible for producing short-chain fatty acids [58], which were upraised in our treatment group.
Genus Lactobacillus, belonging to firmicutes phylum, was enriched in the treatment group (P = 0.05) as in the previous study on several probiotic species [39, 59]. They exert a positive effect on the host health by maintaining immune homeostasis, improving gastrointestinal barrier function, and suppressing pro-inflammatory cytokines [56]. Some species under Lactobacillus genus also can produce several inhibitory substances, including H2O2, that may limit the growth of pathogens to protect intestinal mucosa by strengthening toxic oxidation [60]. In our study, the production of H2O2 was increased for three weeks. This can be another reason for the decrease in species richness in the treatment group. Similarly, Lactobacillus also can reduce the production of endotoxins, LPS [61]. Turicibacter was found to be higher in the encapsulated group. Turicibacter metabolism and its association with the host animal are not well studied, but it has a remarkable action for reducing metabolic stress and has an anti-inflammatory and anti-obesity effect [62]. Furthermore, we observed a significant reduction in genus romboustia, which is considered to be present in a healthy body and maintain the health status of the host. But, it is also reported as a possible marker of gut dysbiosis [63].
Lactobacillus murinus relatively and significantly increased in the treatment group. Lebovitz and Theus, 2019 has reported the applications of L.murinus from the various studies [64] and outlined that it has a beneficial effect on the host, including antimicrobial production, antagonist against pathogens, intestinal barrier, and can be developed as a potential probiotic. Bacteroides thetaiotaomicron helps in Carbohydrate metabolism, lipid metabolism, and enervates the production of pro-inflammatory cytokines and finally helps to strengthen the host-microbiome ecosystem [5]. This species was enriched in our study.
As per the function prediction, the pathway associated with the antimicrobial resistance gene was increased. Biosynthesis of antibiotics may be the reason for the enrichment of the antimicrobial resistance gene. Here, the pathway related to the synthesis of ansamycins (antibiotic) was also enriched. So, further study is necessary to find its validity on commercial production of livestock or poultry for the long run with the different doses as this may develop a negative influence on gut health. Pathway associated with lipoic acid metabolism may elevate host antioxidant properties and anti-inflammation [65].
Epithelial cell of the intestine establishes a physical and chemical barrier for preventing antagonism between immune cells of host and gut microbes to protect the mucosa from inflammation [1]. SIgA is an abundant antibody class found in the intestinal lumen, illustrated as the first line of defense to protect intestinal epithelium from pathogens and enterotoxins and has a key role in immune protection [66]. The result in this study showed that the SIgA quantity was upraised in feces of mice. Similarly, we observed a rise in mucin content and MUC-2 expression after the administration of encapsulated L. paracasei. Mucin fundamentally helps in epithelium lubrication and involved in immune functions. Furthermore, MUC2 covers the intestinal tract and safeguard the GI tract from pathogens attacks [1, 67, 68]. Fecal albumin is another index for measuring intestinal permeability, which estimates the flow from the blood to the lumen. A healthy intestinal barrier checks the overflow of albumin in the intestinal space. A higher value of albumin indicates the disruption of the intestinal barrier [69]. As expected, fecal albumin quantity was reduced in our study, which is a good sign for a healthy intestinal epithelium layer. Cytokines have the function to balance the intestinal immunity in the host. Intestinal Infections lead to inflammation that raises several pro-inflammatory factors [70]. Li et al., 2016 had described that supplementation of Lactobacillus acidophilus against E.coli decreased the secretion of pro-inflammatory cytokines and increased the secretion of anti-inflammatory cytokines compared to control, indicating probiotics can act as an anti-inflammatory supplement [71]. In our current study, the supplementation of encapsulated L. paracasei decreased the expression of TNF-α, IL-1β, IL-6 and IL-8 and increased the expression of IL-10 in different intestinal tissues. Our results are in line with the earlier studies with other probiotics [37, 58, 72]. Thus, we can suggest that encapsulated L. paracasei can show anti-inflammatory effects and protect the gut. But, interestingly, there was a difference in the mRNA expression within a tissue. So, further studies are needed to draw a specific conclusion from this.
Higher production of reactive oxygen species (ROS) than antioxidants leads to oxygen stress in host health. Due to such imbalance, there is a disturbance in a cell leading to damage of DNA, lipids, and proteins. During ROS production, several antioxidant enzymes like superoxide dismutase (SOD) and glutathione peroxidase (GPx) defend against oxidative stress for balancing the system. However, MDA activity is increased during oxidative damage [73]. In the earlier studies, different strain of probiotics including, Lactobacillus plantarum KFY02 [58], Bacillus velezensis, and Bacillus subtilis [37], improved the antioxidant level of animals by increasing antioxidant enzymes. In this study, the quantity of TAOC and GSH-Px were increased while MDA level was decreased, which concludes our encapsulated probiotics can enhance the antioxidant capacity of animals.
{kind=link}