3.1 Identification of PIEZO1 expression level in piezo1 KO zebrafish
As the basis of following phenotypic experiment, we first confirmed the decrease of PIEZO1 expression level at both transcription and translation levels in homozygous piezo1 KO zebrafish. Furthermore, the data in transcription level from wild, heterozygous and homozygous KO strains showed that piezo1 gene has a certain quantitative effect.
3.2 Phenotype identification of piezo1 KO zebrafish in different development level
To compare the phenotypic changes of piezo1 KO homozygous zebrafish heart at different developmental stages, 48hpf and 72hpf fish larvae and adult fish at 7-8 month were selected for phenotypic analysis. From the perspective of morphology analysis, no sign of clear heart development abnormity was identified in fish larvae at 48hpf though certain individual differences were observed (Figure 4a). However, the heart of 72hpf fish larvae showed increased heart size and volume, shown through both the diameter and the depth of the developing heart, and with certain degrees hydropericardium. Then, we characterized 72hpf juvenile fish at which stage clear heartbeat can be observed and found that the heart rate of the homozygous mutants experienced a major decline (Figure 4c). Decrease of blood-pumping ability of the heart was also observed (Data not shown). The adult mutants experienced phenotypic transformation of ventricular enlargement, shown through the passivated ventricular contour and lack of apparent apex (Fig 4d), which was reminiscent of the similarly enlarged heart at 72hpf. This morphology change was also confirmed through HE staining. Additionally, analysis of cell species showed an increase in immune cell infiltration among mutant cardiac cells (Fig 3g-3i).
3.3 Expression level of heart development key genes in piezo1 KO zebrafish at different development stages
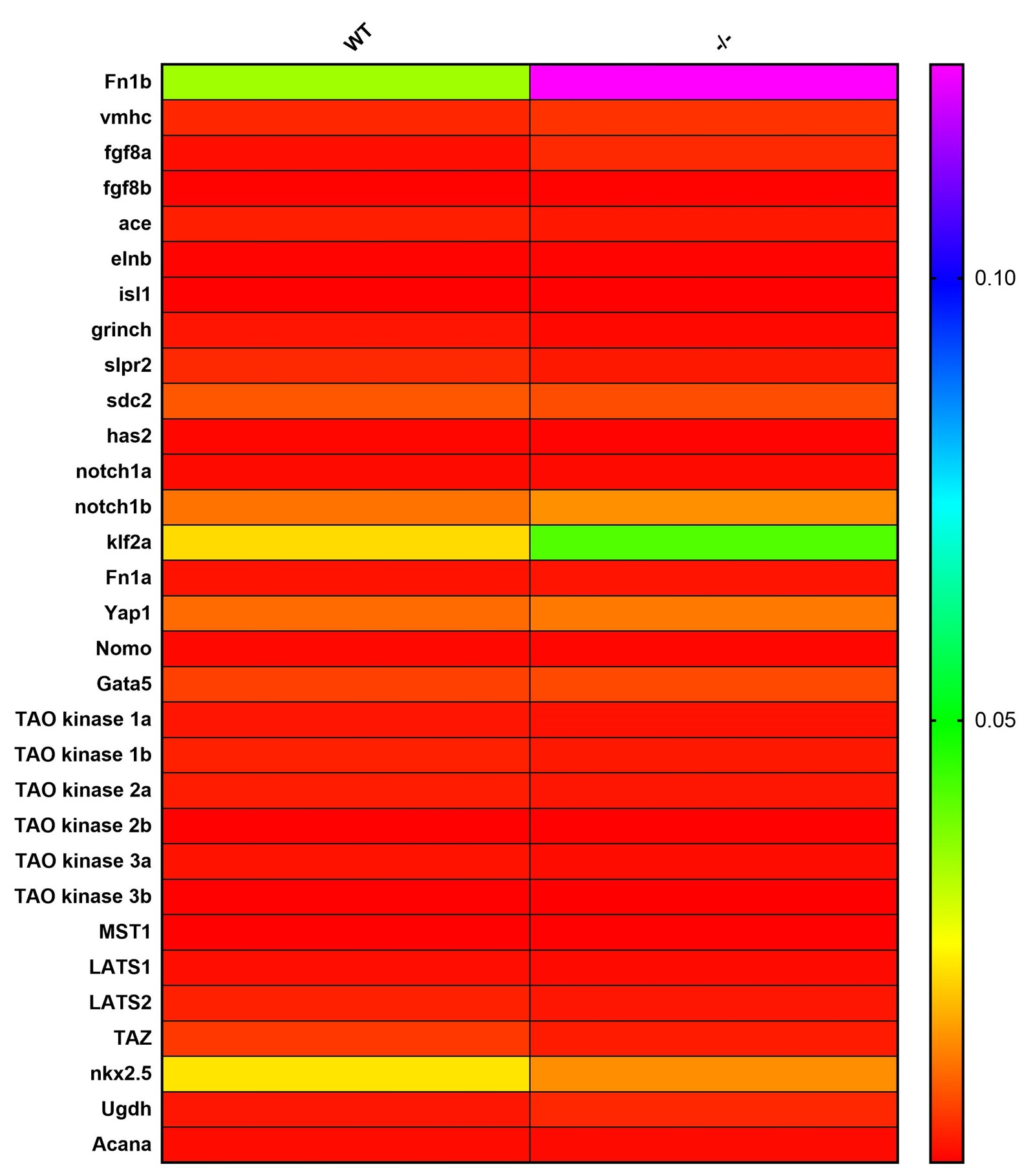
In this study, we chose genes related to the development and migration of embryonic central myocardial cell (grinch, s1p2, sdc2), genes related to the differentiation of myocardial cell (gata5, hand2), genes related to the development of artery (ace/gfg8), vein (Islet-1), arterial glands (elastin b), aortic valve (Notch1), atrioventricular valve(Ugdh), vascular endothelial cell (Fn1, klf2a), ventricular myosin (vmhc) and extracellular matrix of cardiac cells (has2, AcanA, elastin b)[21]. All the genes were detected in wildtype zebrafish and Piezo1 KO homozygous zebrafish at different stages of development. In 48-hour-embryo, the expression level of mutant AcanA Genes showed an obvious up-regulation (Fig 4a). However, the expression of level of these gene did not show any significant changes in whole adult zebrafish lysates (Supplementary Fig 1), but the expression level of hand2 gene showed a significant decrease in dissected hearts of adult zebrafish. Since hand2 is a key factor involved in the differentiation of cardiac muscle cells, this expression will further investigate the changes in the expression level of potential upstream pathway of hand2. Previous research has shown that nkx2.5 plays an important role in the signal transduction in cardiac cells, as the upstream or parallel factor of hand2[21]. Additionally, studies have shown that Hippo pathway can regulate the expression of BMP and hand2 signals through LATS1/2, thus influencing the differentiation of cardiac cells[22]. These two possible upstream pathways were selected for second sound of expression level examination, but no significant transcriptional changes were found (Fig 4c,4d).
3.4 Identification of differentially expressed genes in hearts ofpiezo1-/- zebrafish
To study transcriptomic changes, piezo1-/- heart RNA sequencing was conducted. It was indicated that piezo1 knockout leads to 818 differentially expressed genes (DEGs), in which 535 were upregulated (Fig. 5a). Among these, we concentrated on several DEGs that are related to heart functions, namely upregulated shox2, dsc2l, hbegfb, tfpi2 and downregulated smyd1b. Combined with GO annotations, 5 more genes that were not regarded as DGEs but related to heart functions were selected out (Fig. 5b), including itga4, ptpn21, cenpf, uqcrb and fbxw7. Altogether, the expression changes of these 10 genes were examined through qPCR (Fig. 5c) and showed consistent expression changes in RNA-seq data: shox2, cenpf, itga4, dsc2l and tfpi2 were upregulated, whereas smyd1b, uqcrb, fbxw7 and ptpn21 were downregulated. These heart-function-related genes mainly involve in 3 pathways: heart development enriched by shox2, tfpi2, dsc2l, cenpf, ptpn21 and smyd1b, heart contraction enriched by shox2, smyd1b and hbegfb, and vasculature development enriched by itga4, fbxw7, uqcrb and cenpf.
GO enrichment analysis showed top terms (Fig. 5d, top) in biological process (phosphatidylcholine-sterol O-acyltransferase activator activity, cholesterol binding, and cholesterol transporter activity), cellular component (nucleosome, extracellular space and extracellular region) and molecular function. To further explore the potential pathways DGEs participated in, GO enrichment analysis revealed that DGEs were also closely related to immune responses such as defense response against bacterium, innate immune response and inflammatory response, as well as cholesterol or lipid metabolism including HDL particle assembly, positive regulation of cholesterol esterification and VLDL particle remodeling (Fig. 5d, down).
{kind=link}