CircRNA expression patterns in human OS and chondroma tissues
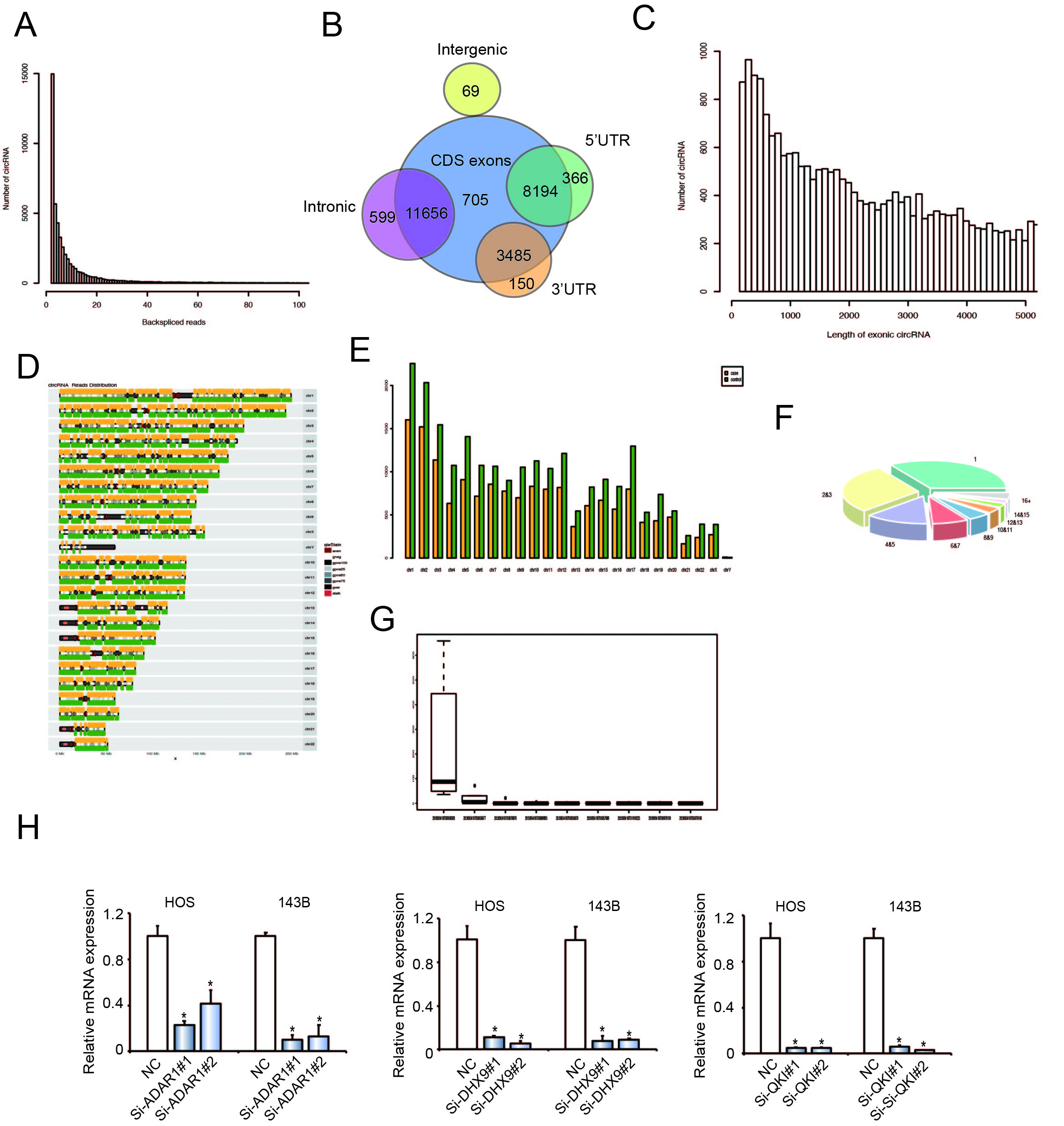
To generate a circRNA-profiling database, we performed circRNA deep sequencing of ribosomal RNA-depleted total RNA from clinical OS and chondroma tissues. RNA from three human osteoblastic osteosarcoma samples and three chondroma tissues were sequenced using an Illumina Hiseq X Ten. The reads obtained were mapped to reference ribosomal RNA (Bowtie2, http://bow tie-bio.sourceforge.net/bowtie2/) and to a reference genome (TopHat2, http://ccb.jhu.edu/software/tophat/) 14,15. Twenty bases from either end of the unmapped reads were extracted and aligned to the reference genome, to identify unique anchor positions within the splice sites. Anchor reads that aligned in the reverse orientation (head-to-tail) indicated circRNA splicing and were then subjected to find_circ (https://omictools.com/find- circ-tool) to identify the circRNAs 16. A candidate circRNA was identified if it was supported by at least two unique back-spliced reads in one sample. A total of 25224 circRNAs were identified using this approach (Supplementary Figure S1A) 17. We next annotated the identified candidates using the RefSeq database 18. Most of the circRNAs originated from protein-coding exons, while others aligned with introns, 5′-UTRs and 3′-UTRs (Supplementary Figure S1B). The majority of identified circRNAs were less than 2000 nucleotides (nt) in length (Supplementary Figure S1C). The chromosomal distribution of identified circRNAs showed no obvious differences between the OS and chondroma groups, while the total expression of circRNAs in the OS group was downregulated (Supplementary Figure S1D and E). Analysis of the number of circRNAs from host genes revealed that one gene could produce multiple circRNAs (Fig. 1F), which is consistent with previous reports19. We further investigated the abundance of circRNAs within one gene locus (n¼3,687). The thick line indicates the median, ends of the boxes define the 25th and 75th centiles and bars define the 5th and 95th centiles. The top five expressed circRNAs were presented in Supplementary Figure S1G. This result indicated that there is often a predominantly expressed circRNA isoform from one gene locus. The OS and chondroma groups displayed differential circRNA expression patterns (Fig. 1A). We focused on candidates that had the greatest differential expression between OS and chondroma groups, then matched them with circbase (http://www.circbase.org/. Among these specific candidates, novel_circ_0064644, which is formed by circularization of exon 7 to exon 10 of RBMS3, attracted our attention.
Identification Of Circrbms3 As A Circrna
To verify that exons 7 to 10 of RBMS3 form an endogenous circRNA, we designed convergent and divergent primers that specifically amplified the canonical or back-spliced forms of RBMS3 (Fig. 1B). Using cDNA and genomic DNA (gDNA) from HOS and 143B cell lines as templates, circRBMS3 could only be amplified using divergent primers, and no amplification product was observed from gDNA (Fig. 1B). By using reverse transcriptase-real-time polymerase chain reaction (RT-qPCR), we further confirmed that circRBMS3 was resistant to RNase R, while RBMS3 mRNA levels were significantly reduced following RNase R treatment (Fig. 1C).
To investigate the function of circRBMS3 in OS development, we assayed the expression level of circRBMS3 in 12 pairs of chondroma and OS tissues, respectively, as well as in OS cell lines. Taking advantage of RT-qPCR and chromogenic in situ hybridization (CISH), we observed, consistent with the RNA-seq analysis, that circRBMS3 expression was higher in OS than in chondroma tissues or normal cells (Fig. 1D-F). Next, Sanger sequencing was used to confirm the circRBMS3 junction (Fig. 1G). Endogenous circRBMS3 was further revealed using a junction-specific probe in northern blotting (Fig. 1H). To establish the cellular localization of circRBMS3, we conducted fluorescence in situ hybridization (FISH) analysis. The junction probe detected abundant cytoplasmic circRBMS3 expression in HOS cells. Additionally, qPCR analysis from different cell fractions confirmed that circRBMS3 is predominantly located in the cytoplasm (Fig. 1I, lower panel).
The expression of circRBMS3 in OS can be regulated by ADAR1
It has been previously reported that RBMS3 is a novel tumor suppressor gene, in esophageal squamous cell carcinoma (ESCC) 20, breast cancer 21, nasopharyngeal cancer (NPC) 22 and lung squamous cell carcinoma (LSCC)23. Although circRBMS3 was significantly upregulated in OS cell lines, the protein levels of RBMS3 were downregulated (Fig. 2I). This finding suggests that the higher expression of circRBMS3 in OS is not simply a by-product of splicing, but may be functional.
Next, we explored the reasons for circRBMS3 upregulation in OS, alongside RBMS3 mRNA and RBMS3 protein downregulation (Fig. 1J). As circRNAs could be regulated by RNA-binding proteins 24 post-transcriptionally, we assumed that circRBMS3, but not RBMS3 mRNA, is regulated by certain RNA-binding proteins post-transcriptionally during human OS development. To test this hypothesis, we measured the expression of circRBMS3 in OS cell lines after individually knocking down all two human RNA-binding proteins 25,26,27 reported to broadly regulate the biogenesis of circRNAs, including adenosine deaminase 1 acting on RNA (ADAR1) and DExH-Box Helicase 9 (DHX9). Among these two RNA-binding proteins, DHX9 has been reported to play an important role in OS devepment28,29,30. After knocking down ADAR1(p110), but not DHX9, circRBMS3 was upregulated, while RBMS3 mRNA did not show significant changes (Fig. 1K). Notably, mRNA and protein levels of ADAR1 were downregulated in OS cell lines (Fig. 1L). Taken together, the upregulation of circRBMS3 is, at least partly, caused by the downregulation of ADAR1 in human OS.
Effects of circRBMS3 on OS and other tumor cell lines
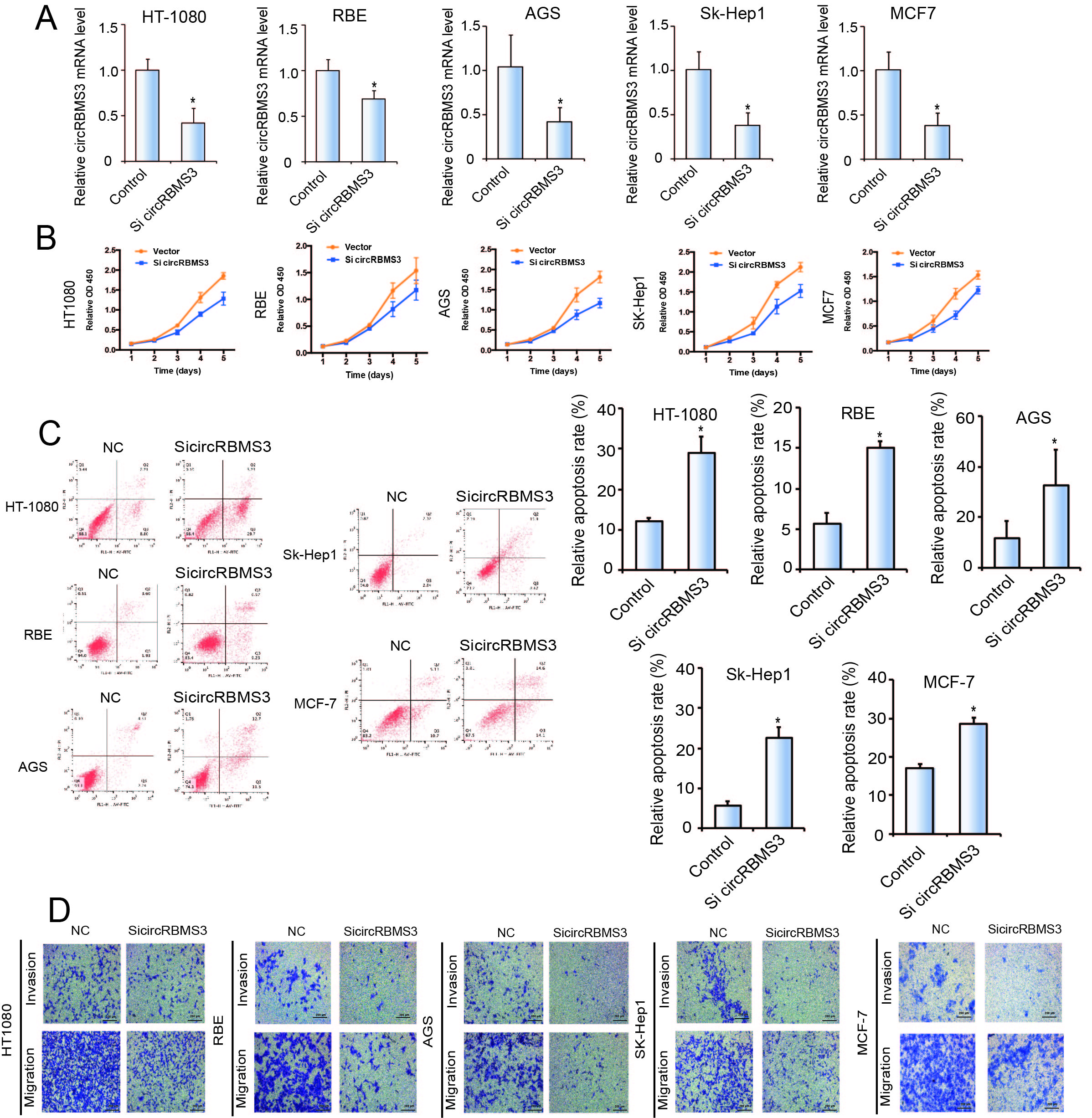
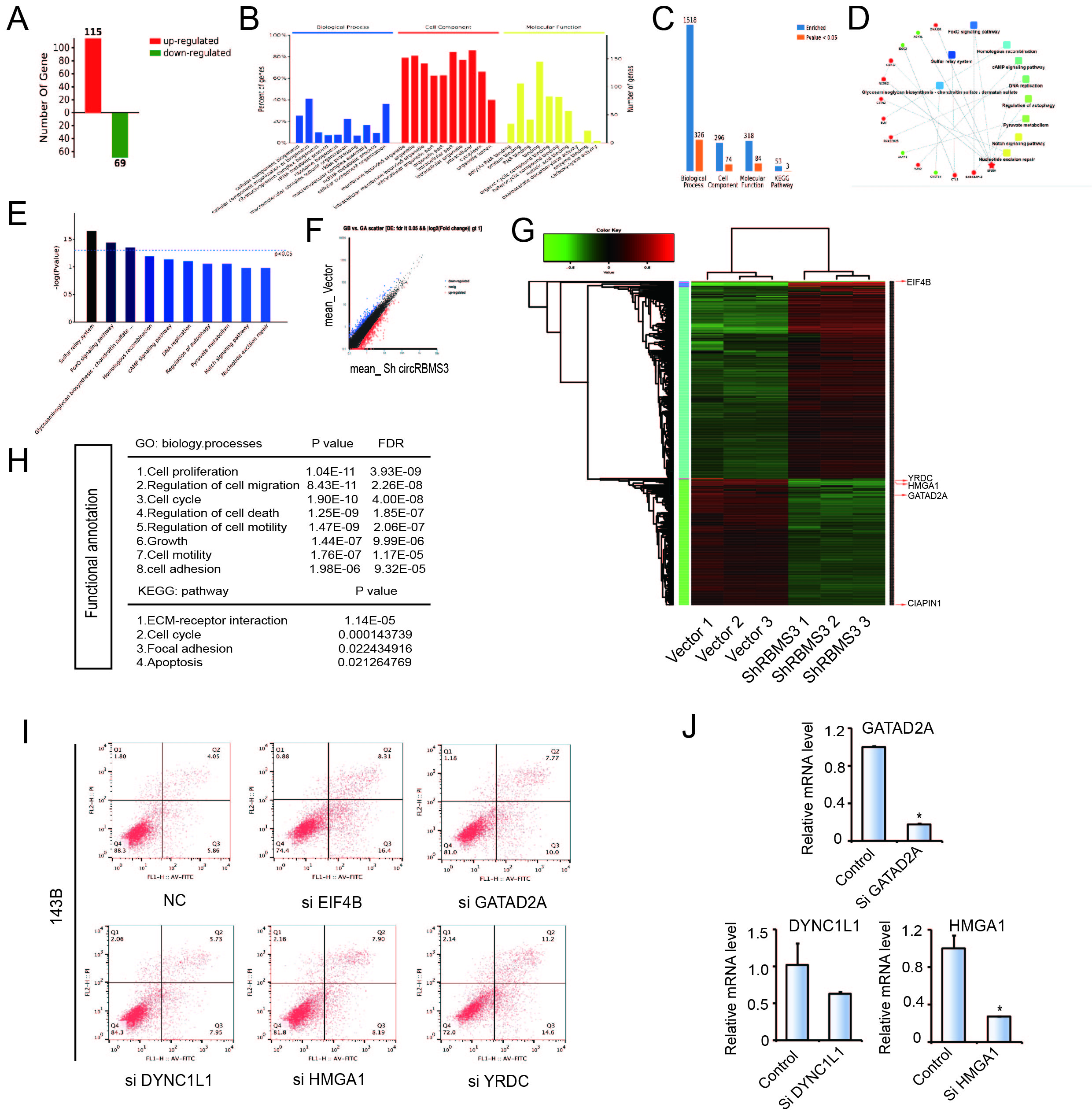
To further explore the biological functions of circRBMS3, we introduced two circRBMS3-knockdown short interfering (si)-RNAs, targeting the junction sites of circRBMS3 into HOS and 143B OS cells. The expression of circRBMS3 was significantly reduced in siRNA-transfected cells (Fig. 2A). Meanwhile, the expression of RBMS3 mRNA showed no apparent change (Fig. 2B). To investigate the function of circRBMS3 in other tumor cells, we investigated the effect of circRBMS3 knockdown in sk-Hep1 (hepatic cellular carcinoma), AGS (gastric cancer), MCF-7 (breast cancer), HT1080 (sarcoma), and RBE (cholangiocarcinoma) cell lines (Supplementary Figure S2A). The proliferation and colony formation abilities of HOS and 143B cells decreased upon circRBMS3 inhibition (Fig. 2C-D), as did that of other tumor cell lines following circRBMS3 knockdown (Supplementary Figure S2B). Flow cytometric analysis was conducted to determine the effect of circRBMS3 knockdown on the apoptosis rate of OS cells and other tumor cells. Notably, inhibition of circRBMS3 augmented cellular apoptosis (Fig. 2E, Supplementary Figure S2C). Consistent with this, a wound-healing assay demonstrated a significant inhibition of cell migration in HOS and 143B cells following circRBMS3 knockdown (Fig. 2F). Moreover, inhibition of circRBMS3 also suppressed migration and invasion of OS cell lines and malignant tumor cell lines in transwell migration and Matrigel invasion assays (Fig. 2G, Supplementary Figure S2D). We also found that transfecting circRBMS3 siRNA did not affect the RBMS3 protein level (Fig. 2H). Knockdown of circRBMS3 did not alter total protein levels (Fig. 2I) or interaction of RBMS3 with its partners (Fig. 2J). Together, these results indicate that circRBMS3 is involved in tumor cell growth, migration, and invasion in vitro.
Molecular Mechanism Of Circrbms3 In Os
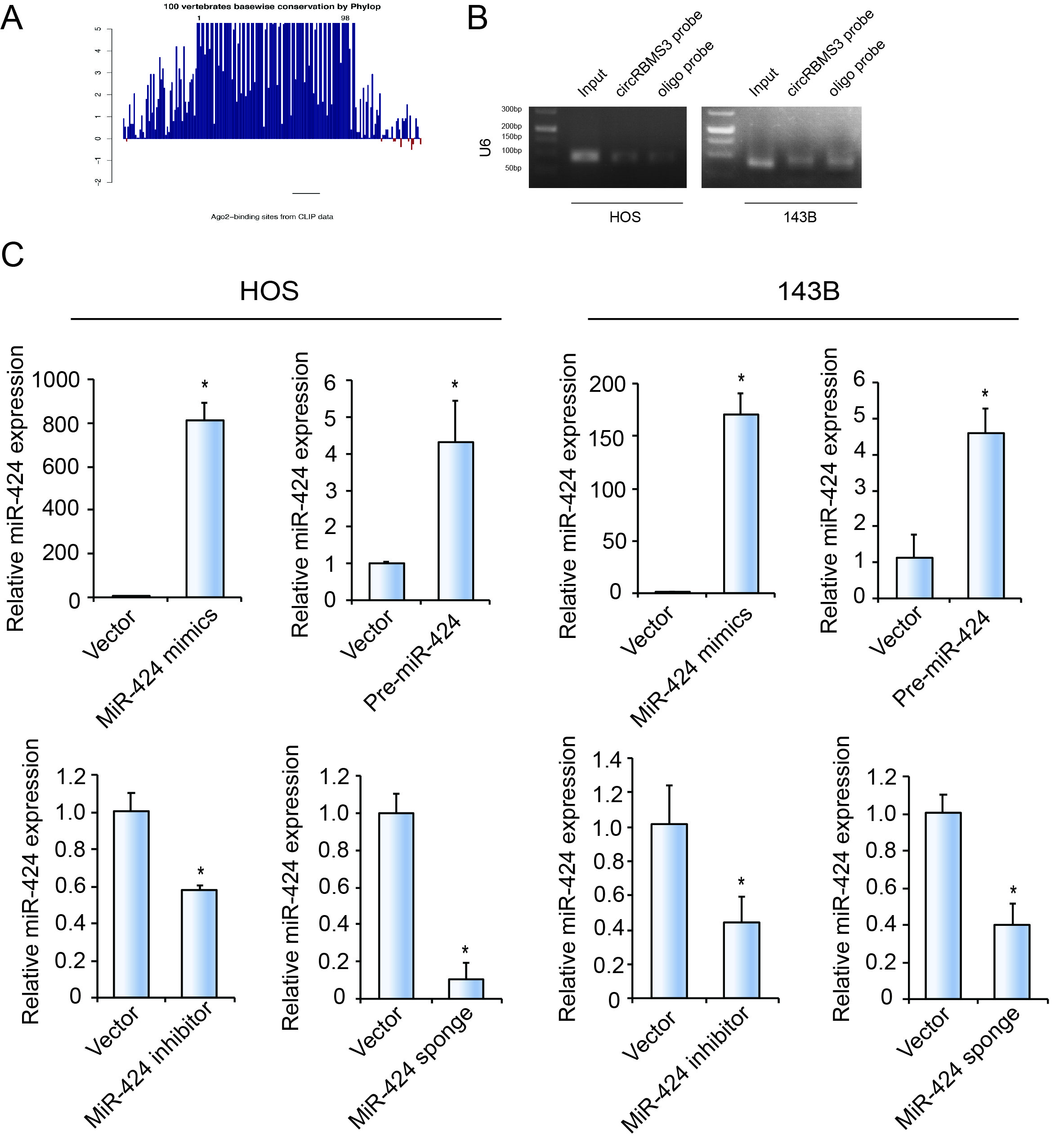
It has been previously reported that circRNA functions as an miRNA sponge in cancer cells8,9. As circRBMS3 is abundant in the cytoplasm, we performed an analysis using freely available AGO2 immunoprecipitation data, including high-throughput sequencing from doRiNA. We observed a high degree of AGO2 occupancy in the circRBMS3 region, which is highly conserved across several vertebrate species (Supplementary Figure S3A). To validate this result, we conducted RNA immunoprecipitation for AGO2 in 143B cells and found that endogenous circRBMS3 pulled-down from AGO2 antibodies was specifically enriched by RT-qPCR analysis when compared to circRBMS3 knockdown cells (Fig. 3A). To confirm whether circRBMS3 could aggregate miRNAs in OS cells, we selected 11 candidate miRNAs by overlapping the prediction results of miRNA recognition elements in the circRBMS3 sequence, using miRanda (Score Threshold > 140), Targetscan(at least 6mer binding site), and RNAhybrid (Fig. 3B). Next, we investigated whether circRBMS3 could directly bind these candidate miRNAs. A biotin-labeled circRBMS3 probe was designed and verified in OS cell lines, and the pull-down efficiency was established (Fig. 3C and D, Supplementary Figure S3B). The miRNAs were extracted following the pull-down assay, and the levels of 11 candidate miRNAs were determined by RT-qPCR. As shown in Fig. 4E, the miR-15a/497/424 cluster was the only one abundantly pulled down by circRBMS3 in both HOS and 143B cells. Because circRBMS3 contained miR-15a/497/424 binding sites (Fig. 3F), we transfected miR-15a/497/424 mimics into HEK-293T cells and observed the insertion of the circRBMS3 binding sequence in the 3′ untranslated region (3’-UTR) downregulated luciferase activity (Fig. 4G). Compared with controls, miR-424 decreased luciferase activity to the greatest extent, (~ 45%) (Fig. 3G). We then mutated the binding sites and transfection of the 3 miRNAs had no significant effect on luciferase activity when the corresponding target sites were mutated in the luciferase reporter (Fig. 3G). We then chose miR-424 for further investigation. FISH assays revealed that circRBMS3 interacts with miR-424 in the cytoplasm of OS cells (Fig. 3H). These results suggested that circRBMS3 functions as a sponge for miR-424.
MiR-424 inhibits OS cell migration, invasion, and proliferation in vitro
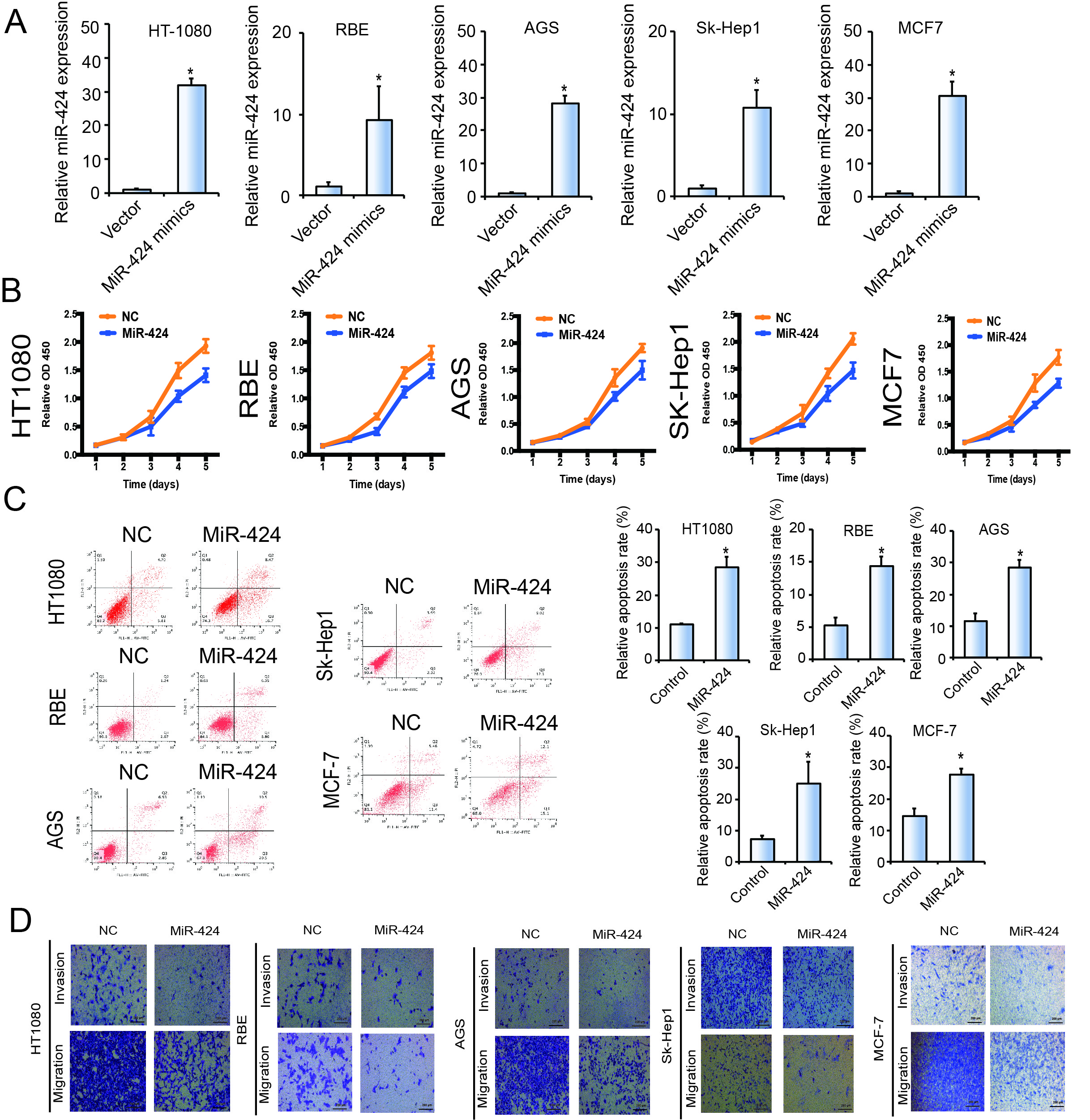
Using qPCR and FISH, we observed miR-424 downregulation in human OS samples compared with chondroma tissues (Fig. 4A and B). MiR-424 was also expressed at low levels in OS cells compared with hFOB1.19 cells (Fig. 4C). We then investigated whether miR-424 has a tumor repressor role during OS cell proliferation. Pre-miR-424 or miR-424 sponge were transfected into HOS and 143B cell lines, as well as the control vector. Expression of miR-424 was verified by RT-qPCR (supplementary Figure S3C & FigureS 4A). Results of CCK-8 and colony formation assays showed that overexpression of miR-424 in OS cells results in significant inhibition of cell proliferation (Fig. 4D and E, supplementary Figure S4B). In contrast, the proliferation rates of OS cells transfected with miR-424 sponge were significantly higher than those transfected with the control vector (Fig. 4D). Furthermore, we demonstrated that ectopic expression of miR-424 clearly increases the apoptosis rate 48 h after transfection of miR-424 mimics (Fig. 4F, supplementary Figure S4C). In addition, the wound healing assay (Fig. 4G) and Transwell assay (Fig. 4H, supplementary Figure S4D) showed that overexpression of miR-424 inhibits the migration and invasion of OS cells. Furthermore, Kaplan-Meier survival curves of the TCGA sarcoma dataset showed that patients with high expression (cut-off by the median gene expression value) of miR-424 had an elevated 10-year overall survival rate (Fig. 4I). These results confirmed that miR-424 reduced the proliferation, migration and invasion ability of osteosarcoma cells.
EIF4B and YRDC are direct targets of miR-424
Bioinformatics analysis was used to search for potential regulatory targets of miR-424. We got 1287 potential target of miR-424 from Targetscan (www.targetscan.org) with the standard “at least 6mer binding sites”, 270 circRBMS3-regulating genes from RNA sequence and 190 circRBMS3-regulating proteins from Mass Spec analysis. We selected 5 candidate genes by overlapping the prediction results of gene recognition elements using mRNA sequencing, Targetscan, and Mass Spec analysis (Fig. 5A-D, Supplementary Figure S5A-H). Among these genes, EIF4B and YRDC, whose inhibition leads to the highest apoptosis rates in OS, were selected as targets for further analysis (Supplementary Figure S5I and J). RT-qPCR and immunohistochemistry demonstrated that EIF4B and YRDC were overexpressed in OS tissues compared with chondroma tissues (Fig. 5E-F). In addition, Kaplan-Meier survival curves from the TCGA sarcoma dataset showed that patients with high expression (cut-off by the median gene expression value) of EIF4B or YRDC had a lower 10-year overall survival rate (Fig. 5G).
To verify whether EIF4B and YRDC are direct targets of miR-424, we constructed 3’UTR sensors and co-transfected HEK-293T cells with miR-424 mimic or NC (negative control). We observed reduced luciferase activity of EIF4B and YRDC 3’ UTR in the presence of miR-424 (Figs. 5H and I). To verify target specificity, we generated mutated forms of the EIF4B and YRDC 3′-UTR, in which the miR-424-binding site was abolished (Fig. 5H). Co-transfection of miR-424 mimics alongside the mutant construct abrogated the decrease in wild-type 3’-UTR luciferase activity, indicating that miR-424 specifically regulates EIF4B and YRDC expression (Fig. 5I).
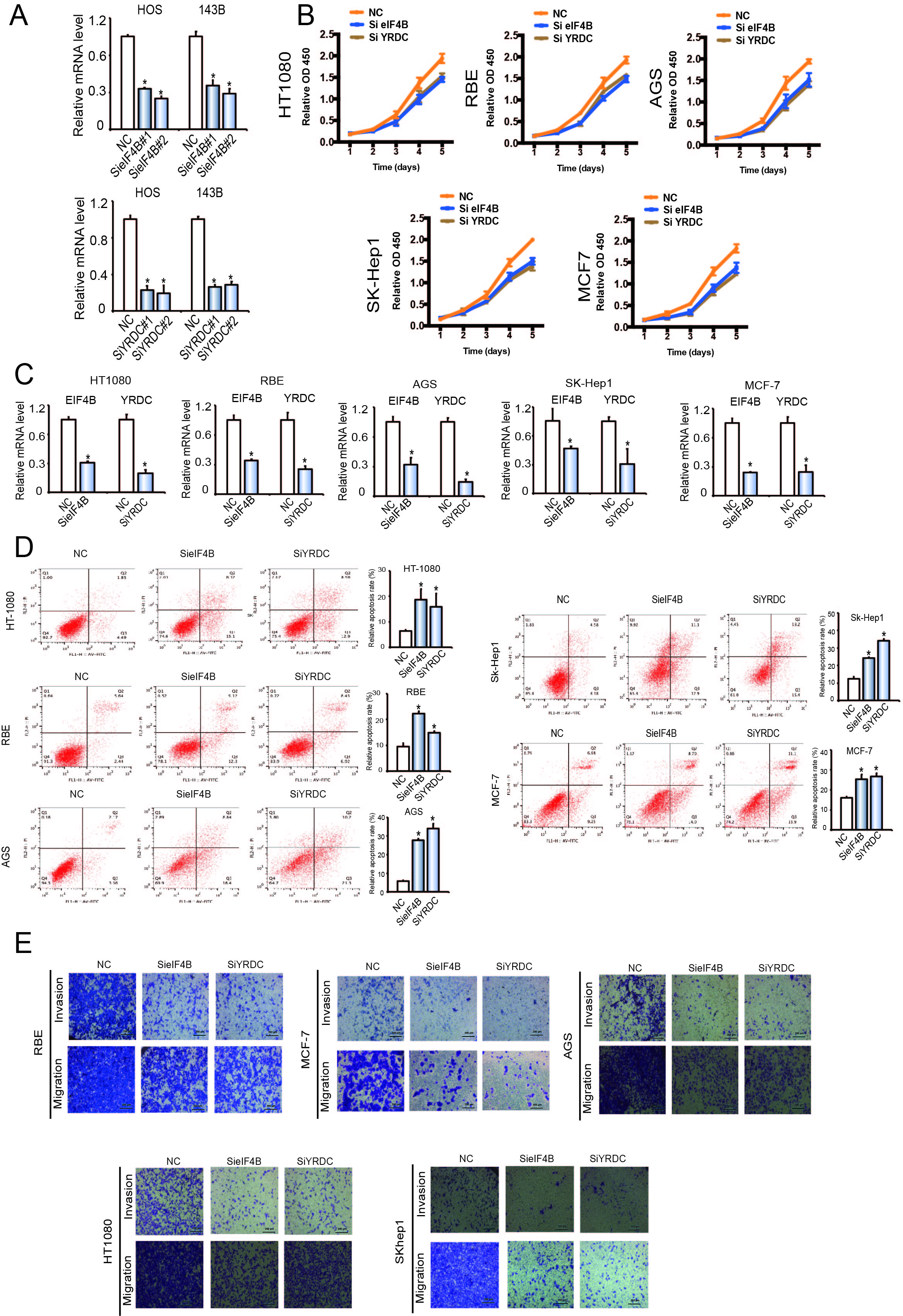
To determine whether miR-424 could affect the expression of eIF4B and YRDC, HOS and 143B cells were transfected with miR-424 mimics, inhibitors, or respective controls. The results of immunofluorescence (Fig. 5J) and western blotting analyses (Fig. 5K) showed that miR-424 mimics markedly suppressed eIF4B and YRDC protein levels in OS cells, while the miR-424 inhibitor clearly promoted eIF4B and YRDC protein expression. Moreover, following transfection, mRNA and protein levels of eIF4B and YRDC were similarly downregulated (Fig. 5L). In summary, these results strongly suggest that miR-424 directly regulates eIF4B and YRDC in OS cell lines.
EIF4B and YRDC are known as oncogenes in some tumor types, while their function in OS is currently not understood. We explored the effect of eIF4B and YRDC on OS tumorigenesis by CCK-8 and colony formation experiments. There was a significant decrease in proliferation after inhibition of eIF4B and YRDC expression (*P < 0.05; Fig. 6A-B, supplementary Fig. 6A-C). Furthermore, knockdown of eIF4B and YRDC clearly induced higher levels of apoptosis (Fig. 6C, supplementary Fig. 6D, *P < 0.05). In addition, the wound healing (Fig. 6D) and transwell assays (Fig. 6E, supplementary Fig. 6E) showed that inhibition of eIF4B and YRDC inhibits the migration and invasion of tumor cells. These results suggested that eIF4B and YRDC are tumorigenic and are direct targets of miR-424.
Knockdown of miR-424 reverses shcircRBMS3-induced attenuation of OS cell proliferation, migration, and invasion
We co-transfected miR-424 sponge and the circRBMS3 knockdown construct into OS cells. Both protein and mRNA expression of eIF4B and YRDC significantly increased in OS cells co-transfected with shcircRBMS3 plasmid and the miR-424 sponge compared with cells transfected with shcircRBMS3 alone (Fig. 7A and B). Immunofluorescence analysis confirmed that the expression of eIF4B and YRDC increased in 143B and HOS cells transfected with shcircRBMS3 and miR-424 sponge together when compared with shcircRBMS3 alone (Fig. 7C). Knockdown of both miR-424 and circRBMS3 resulted in a higher growth rate than the circRBMS3 inhibition group (Fig. 7D). In addition, downregulation of both miR-424 and circRBMS3 promoted colony formation when compared with cells transfected with shcircRBMS3 alone (*P < 0.05 for both; Fig. 7E). Furthermore, apoptotic cell numbers were decreased after treatment with miR-424 sponge and shcircRBMS3 in co-transfected cells compared with shcircRBMS3 transfected cells. (Fig. 7F). Furthermore, wound healing assays (Fig. 7G) and transwell Matrigel TM invasion experiments (Fig. 7H) indicated that OS cells co-transfected with miR-424 sponge and shcircRBMS3 expression constructs demonstrated enhanced invasion and migration capabilities when compared with shcircRBMS3-transfected cells. Strikingly, we observed that inhibition of miR-424 in OS cells significantly strengthened the anchorage-independent growth ability of circRBMS3 knockdown cells (Fig. 7I). Together, these data suggested that circRBMS3 promotes cell migration, invasion, and proliferation via sponging miR-424 and subsequently induces EIF4B and YRDC expression in vitro.
CircRBMS3 functions as miR-424 sponge to promote tumorigenesis in vivo
To explore the effects of circRBMS3 and miR-424 in vivo, 143B cells transfected with circRBMS3-deficient, miR-424 sponge, or vector were injected subcutaneously into nude mice for 35 days and then tumor volumes were measured. Compared with circRBMS3-deficient stable cell, cells lacking both miR-424 and circRBMS3 had a higher tumor growth rate (Fig. 8A and B). Similarly, co-expression of miR-424 inhibition and circRBMS3 knockdown constructs rescued the volume of 143B-derived tumors in vivo (Fig. 8C), compared with the circRBMS3-deficient group alone. We observed the same results for average tumor wet weight across all 3 groups (Fig. 8D).
To further evaluate the antitumor effect of circRBMS3 in vivo, an orthotopic OS model was established by intra-tibial injection of 143B cells. The mice were intra-tibial injected with circRBMS3-deficient, miR-424 sponge, or vector stable cells. As indicated in Fig. 8E and F, in vivo imaging results showed that circRBMS3 inhibited the growth of in situ tumors, while mice with tumors lacking both miR-424 and circRBMS3 had a higher tumor size and posterior limb weight (Figs. 8G).
We next evaluated the relationship between circRBMS3 and two target genes in vivo. CircRBMS3 knockdown reduced the levels of eIF4B and YRDC mRNA (Fig. 9H). This corresponded to a reduction of eIF4B and YRDC expression, as determined by western blot (Fig. 8K). Moreover, inhibition of both miR-424 and circRBMS3 rescued these effect (Fig. 8H-K). As shown in Fig. 8L, circRBMS3 knockdown in tumor tissues resulted in a significant increase of terminal dUTP nick end labeling (TUNEL)-positive cells, whereas the levels of Ki67, eIF4B or YRDC were decreased, while inhibition of miR-424 reversed these effects.
Furthermore, we investigated the effect of circRBMS3 on bone microarchitecture of the tumor-bearing tibia using micro computed tomography (CT) in vivo. As a result, circRBMS3 knockdown significantly reduced the bone destruction (Fig. 8M). Indeed, the trabecular bone volume (BV/TV; from 0.013 to 0.208) was significantly improved after circRBMS3 knockdown (Fig. 8M). The same tendency was observed for the BV (12.3 mm3 in control vs. 21.3 mm3 in treated group), while miR-424 inhibition rescued these effects (Fig. 8M). These results suggested that circRBMS3 acts as a sponge for miR-424, and that miR-424 mediates the tumorigenic function of circRBMS3 in vivo (Fig. 8N).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}