Developing novel TPAP molecular markers for Pita, Pib and Pid2
Pita encodes a cytoplasmic membrane receptor protein containing an NBS domain and a leucine-rich domain (LRD), and its disease-resisting mechanism is due to its detection of the expression product of avirulent gene Avr-Pita in rice blast, thus triggering rapid local defenses (Bryan et al. 2000). Two other useful alleles function in rice blast resistance. One is Pib, which belongs to the NBS-LRR disease-resistant gene group and encodes a protein product comprising one NBS and 17 LRR. The other is Pid2, which encodes a transmembrane receptor protein kinase (RLK) and confers high resistance to the Chinese rice blast strain ZB15 (Wang et al. 1999; Wang et al. 2015). One major task of rice breeders is to use these genes effectively to generate novel highly resistant varieties. Functional molecular markers have been developed for Pita and Pib detection, with two pairs of primers and double PCR amplification needed (Fjellstrom et al. 2004; Wang et al. 2012), while Pid2 allele detection still requires sequencing (Wang et al. 2015). However, the detection primers for Pita, Pib and Pid2 are inefficient for pyramiding multiple genes because the process is time-consuming (Fjellstrom et al. 2004; Kim et al. 2016). Therefore, rapid, simple and low-cost functional molecular markers for Pita, Pib and Pid2 detection are urgently needed.
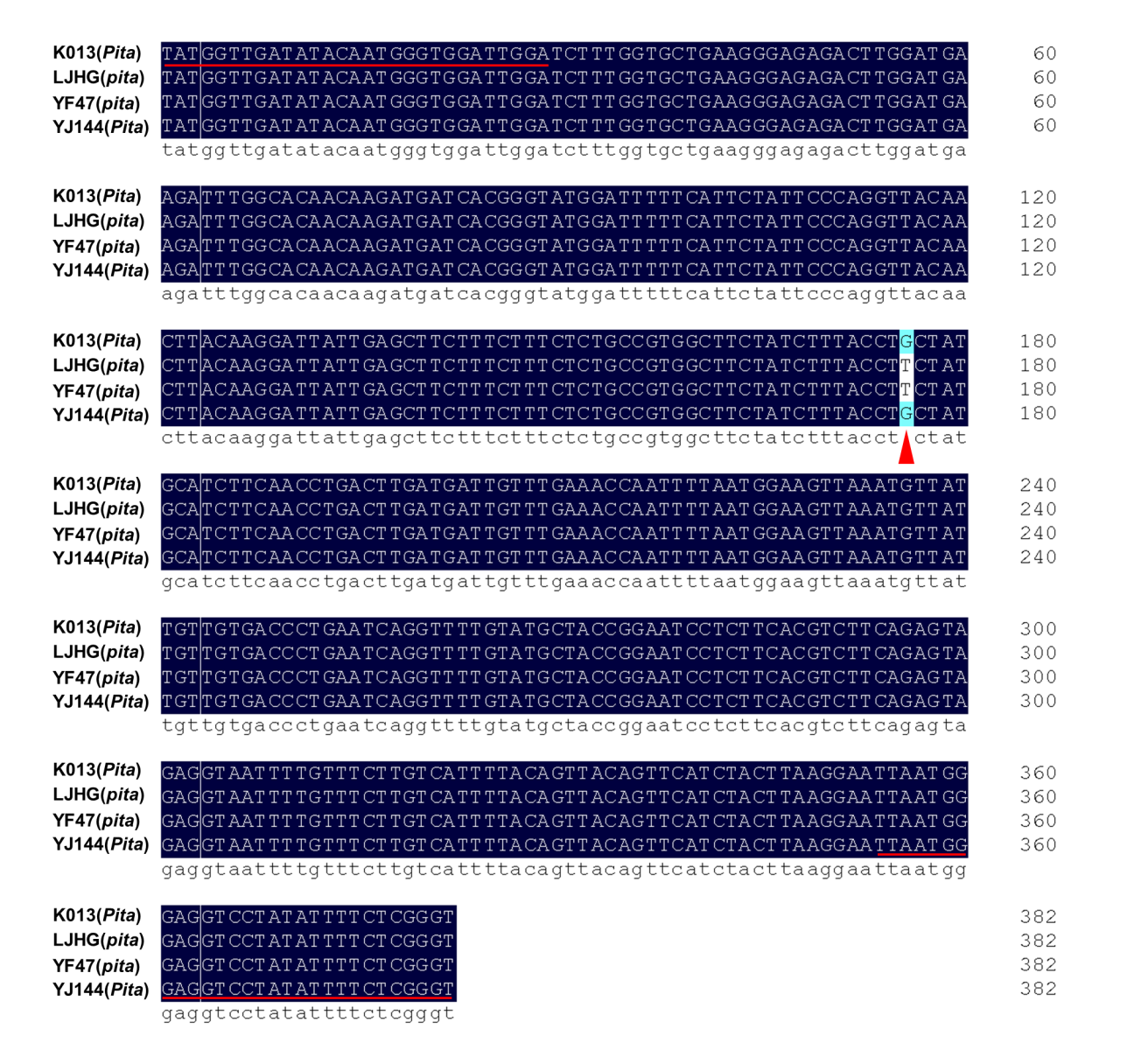
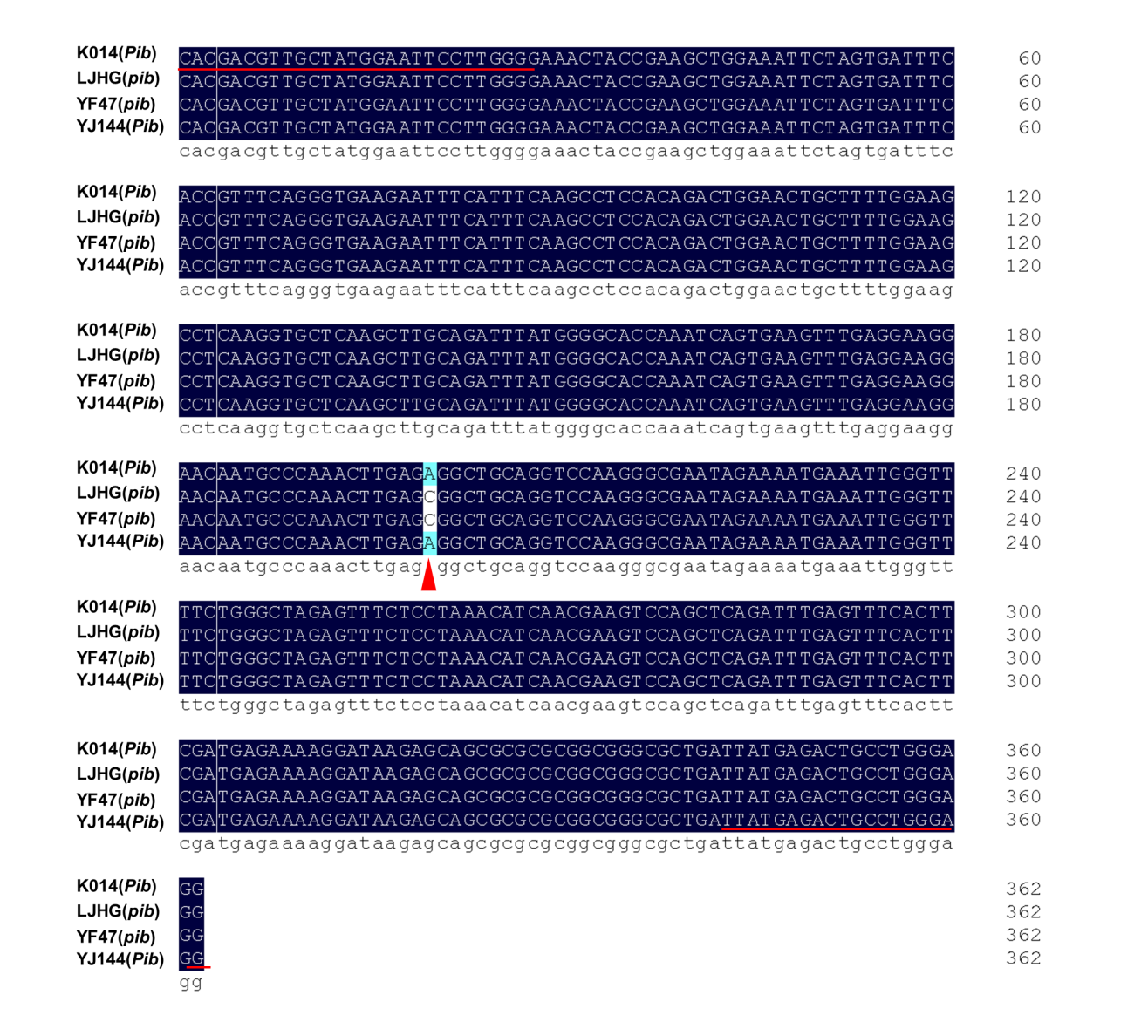
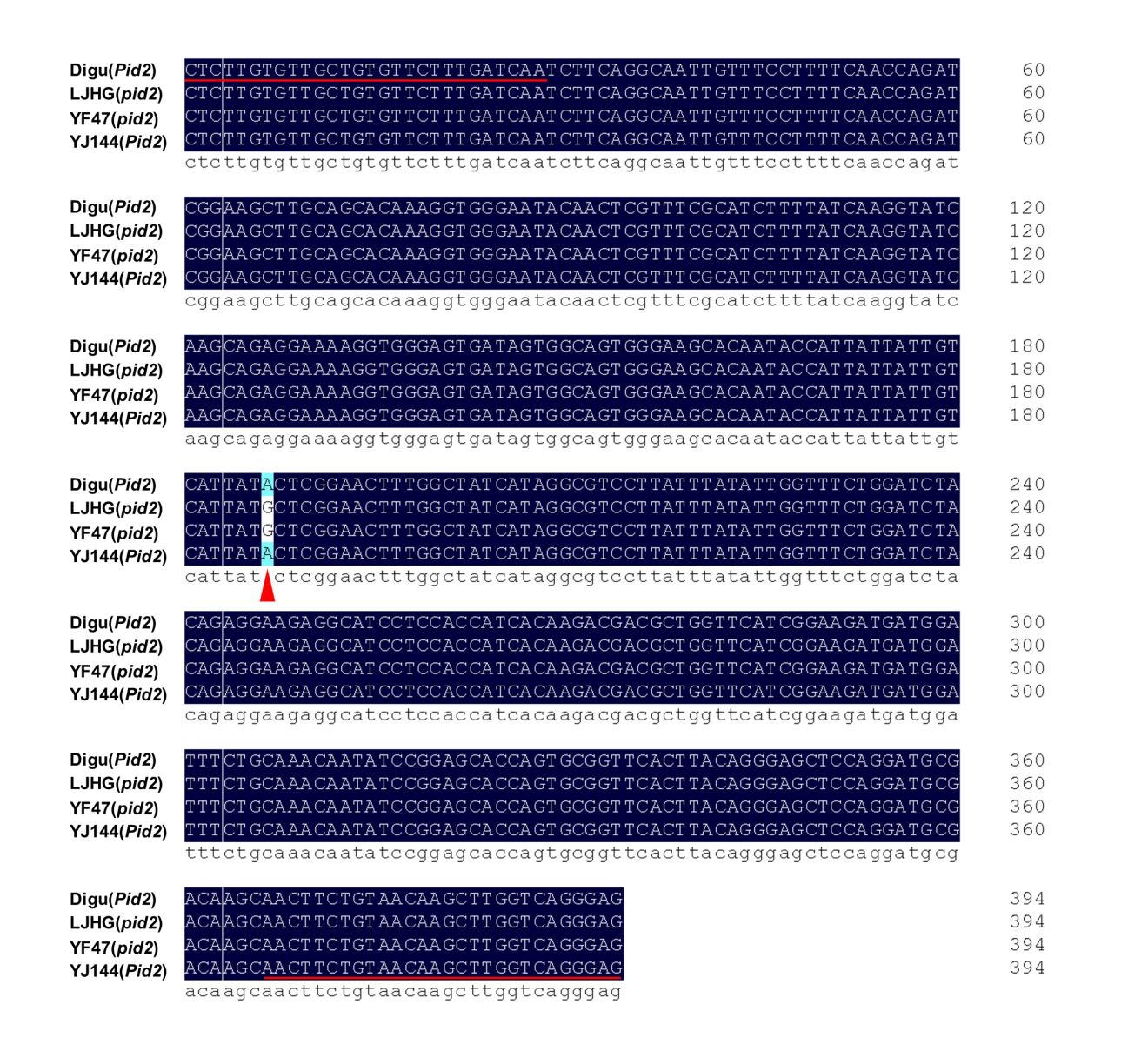
First, based on the functional nucleotide polymorphism sites of Pita/pita, Pib/pib and Pid2/pid2 (LOC_Os12g18360 and LOC_Os06g29810 collected from the MSU database, and Os02g0818450 from RAP-DB), molecular markers were designed for TPAP–Pita, TPAP–Pib and TPAP–Pid2 (Fig. 1A–C) (Wang et al. 1999; Bryan et al. 2000; Wang et al. 2015). Referring to the previous studies, K013, K014 and Digu were chosen as the resistance checks for Pita, Pib and Pid2, respectively, and LJHG was used as the susceptibility check (Wang et al. 2012; Wang et al. 2015). For Pita, the marker TPAP–Pita amplified 230 bp and 201 bp DNA fragments corresponding to the resistance and susceptibility alleles, and a 382 bp fragment was the positive control (Fig. 1D). For Pib, the marker TPAP–Pib amplified 228 bp and 192 bp DNA fragments corresponding to the resistance and susceptibility alleles, and a 362 bp DNA fragment was the positive control (Fig. 1E). For Pid2, the marker TPAP–Pid2 amplified 212 bp and 236 bp DNA fragments corresponding to the resistance and susceptibility alleles, and a 394 bp DNA fragment was the positive control (Fig. 1F). To confirm the precise application of molecular markers, we sequenced the PCR products using outer primers of TPAP–Pita, TPAP–Pib and TPAP–Pid2, and confirmed the accuracy of the above constructed molecular markers (Fig. S1–S3).
Generation and resistance analysis of blast resistance materials using the crosses of YF47 as a receptor parent, and WYJ27 and P135 as donor parents
To investigate the benefit that blast-resistant alleles confer, we studied 91 japonica varieties (or lines) from different provinces: Liaoning, Ningxia, Tianjin, Xinjiang, Jilin and Jiangsu (Table S1). Most of them show similar ecological characteristics and were further used for genotype analysis of Pita, Pib and Pid2. It was found that some cultivars contained one or two functional alleles; however, no one rice variety contained all three (Fig. 1G). Next, considering genetic relationship, plant architecture, rice yield and quality, WYJ27 (with Pita and Pib alleles) and P135 (with the Pid2 allele), were selected as donor parents to improve rice blast resistance of YF47. As a result, a series of rice lines carrying one, two or three resistant genes was identified using multiple strategies of rice breeding, including single, mutual and backcrosses together with MAS (Fig. 2). To accelerate the breeding process further, the anther culture technique was used to generate these genetic materials. A total of 7000 anthers were collected from BC1F1–7 plants at the late uninucleate stage. From these, about 630 embryoids were induced; callus induction was 9%. Finally, 151 green seedlings were successfully generated, giving a plant differentiation rate of 24% (Fig. 2).
To further evaluate the resistance levels of those generated materials, we recorded a leaf and neck blast disease index in both 2019 and 2020 using natural inoculation (Fig. 3). In 2019, at the tillering stage, the leaf blast disease index for No resistance gene Lines (NL, n = 6), Pita (n = 26), Pib (n = 15), Pid2 (n = 22), Pita + Pib (n = 22), Pita + Pid2 (n = 15), Pib + Pid2 (n = 10) and Pita + Pib + Pid2 (n = 5) was recorded as means of 4.1, 2.6, 3.7, 3.8, 1.8, 1.9, 2.8 and 0.4, respectively (Fig. 3A). At the heading stage, the corresponding means were 4.8, 3.4, 4.1, 4.1, 2.5, 2.8, 3.6 and 0.8, respectively (Fig. 3B). The receptor parent YF47 had a leaf blast disease index of 4 and 5 at the tillering and heading stages, respectively. The neck blast severity was recorded as infection percentages of 68.6, 35.7, 42.9, 44.7, 24.8, 28.7, 35.6 and 7.0%, respectively, whereas 68.0% was observed for YF47 (Fig. 3C). In 2020, the disease index increased slightly but showed a similar pattern that of 2019 (Fig. 3D–F). The other two donor parents, WYJ27 and P135, were also used to investigate the disease index. They both showed improved leaf blast disease index and reduced percentage of infection over the control parent YF47 (Fig. 3). Taken together, Pita showed the best improvement against rice blast, while Pib exhibited slightly stronger resistance than Pid2. It is noteworthy that blast resistance increased with the number of functional alleles in the line.
Agronomic traits of YF47 and the improved lines carrying three resistance genes with and without application of fungicides
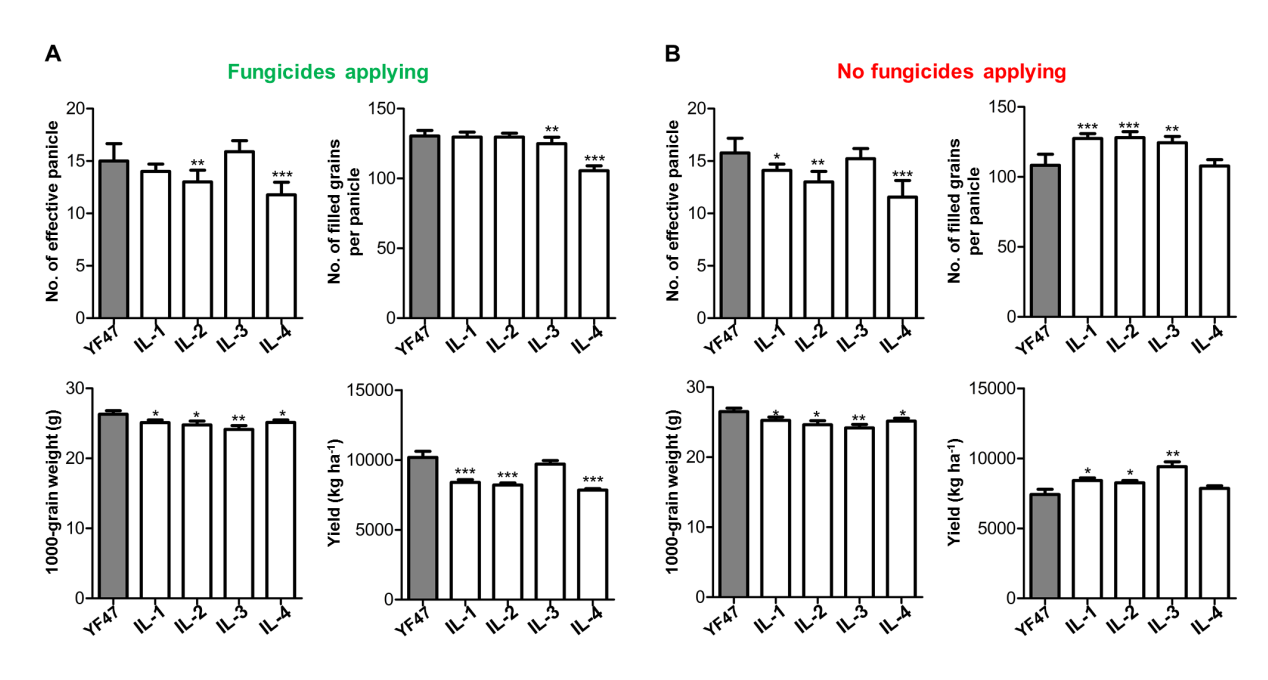
The field performances in Liaoning province were tested for five improved lines IL-1 to IL-5, all with introduced Pita, Pib, and Pid2 alleles with YF47 as a control. Their yield-related traits, including the number of effective panicles, filled grains per panicle, 1000 grain weight and total yield per ha, were investigated with and without the application of fungicides. In 2019, under the application of fungicides, IL-5 showed an increased number of effective panicles, whereas IL-2 and 4 exhibited the opposite outcome. IL-3, 4, and 5 all showed reduced numbers of filled grains per panicle, while the five lines all showed decreased 1000 grain weight. As a result, each of the three lines IL-1, 2, and 4 exhibited a reduction in yield compared to YF47. The other two lines, IL-3 and 5, had yields equal to that of YF47 (Fig. 4A and Fig. S4A). In 2019, without application of fungicides, IL-5 showed an increased number of effective panicles, whereas IL-1, 2, and 4 exhibited the opposite effect. IL-1, 2, 3, and 5 all had an increased number of filled grains per panicle. Also, lines IL-1 to 4 all showed decreased 1000 grain weight. As a result, after rice blast infection, all lines except IL-4 exhibited better yield performance compared to that of YF47 (Fig. 4A and Fig. S4B). Thus IL-3 and 5 are two candidate lines with huge for rice breeding.
To evaluate further the performance of IL-5, with and without application of fungicides, similar experiments were conducted in 2020. We compared the yield-related parameters of YF47 and IL-5. As shown in Fig. 4B, under the application of fungicides, the effective panicle number of IL-5 was significantly higher than that of YF47 (p < 0.01), while the filled grains per panicle and 1000 grain weight were significantly lower than those of YF47 (p < 0.05), resulting in nearly the same yield. However, where fungicides were not applied, the effective panicle number and the filled grains per panicle of IL-5 were both significantly higher than those of YF47 (p < 0.01), while its 1000 grain weight was significantly lower (p < 0.05), resulting in higher yield for IL-5 (p < 0.001).
YJ144, a high blast resistance rice line, exhibited increased yield traits in the Liaoning province regional variety comparison tests
Of five improved lines each containing three rice blast resistance genes, IL-5 had the minimum disease index for leaf blast and neck blast, and showed increased yield both in 2019 and 2020 (Fig. 3–4 and Fig. S4). We named this line YJ144. Then we conducted phenotypic and morphological analyses of YF47 and YJ144. As shown in Fig. 5A–B, YJ144 showed similar plant architecture and panicle morphological architecture compared to YF47. Moreover, YJ144 had higher rice blast resistance (Fig. 5C–D), consistent with their relative leaf and neck blast disease index, as indicated in Fig. 3. To confirm that the three functional alleles Pita, Pib and Pid2 had been successfully introduced, we sequenced the appropriate functional sites of these genes within YF47 and YJ144. The results confirmed that YJ144 contained these novel functional sites, as shown in Fig. S1-S3.
To examine the performance of YJ144 in field trials, in 2019 and 2020 it was entered in the regional variety comparison tests in Liaoning province. Liaojing 9 (LJ9) is an officially designated control variety with medium yield used in regional comparison tests in Liaoning province. Its planting has covered nearly 0.6 million hectares over the past 30 years (https://www.ricedata.cn/variety/varis/600590.htm, accessed 25 July 2020). This trial encompassed tests in six Liaoning cities: Tieling (TL), Shenyang (SY), Liaozhong (LZ), Linghai (LH), Dandong (DD) and Panjin (PJ). In both 2019 and 2020, YJ144 exhibited significant yield increases of 4.3 to 17.5% at all six sites apart from SY. Yields at SY also increased in 2019 and 2020 but not significantly (Fig. 5E–F). These findings suggest that YJ144 exhibited superior yield capacity and could be used as an important japonica germplasm with both high blast resistance and yield.
{kind=link}
{kind=link}
{kind=link}
{kind=link}