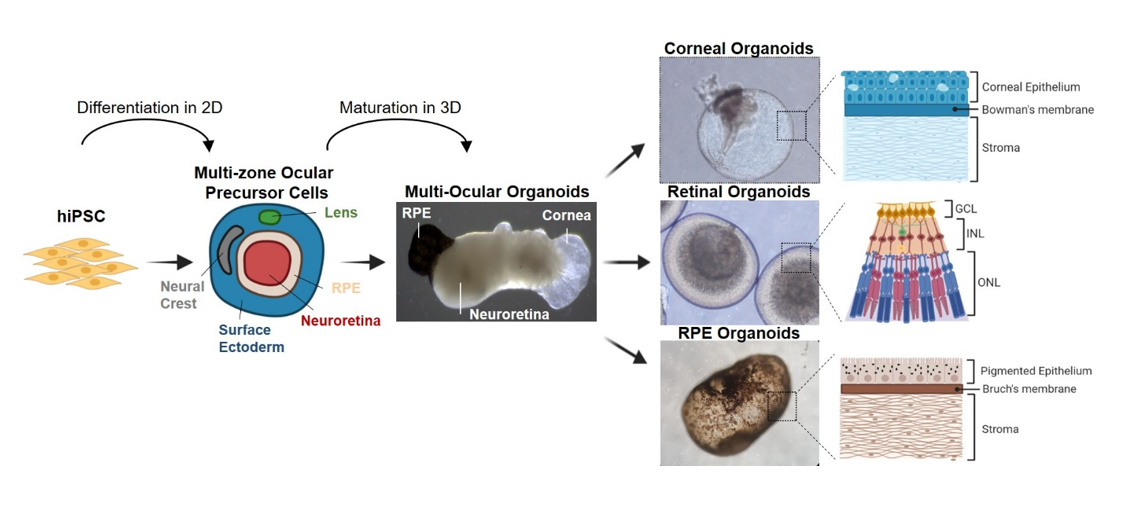
Organoids are 3D cell culture models that closely resemble human embryonic tissues. Emerged by self-organization, organoids mimic mechanisms of normal tissue development, follow typical organ morphogenesis, and display some organ specific functions. This is particularly interesting for studying the eye, which comprises different highly specialized tissues derived from various cell linages, including NE, SE, NC and the periocular mesenchyme (42). While tremendous progress has been made into understanding the highly complex mechanisms involved in eye-field differentiation, current in vitro protocols fail to emulate the spatial cellular organization of the eye. Most differentiation protocols of hiPSC-derived ocular cells and organoids focus on a single -tissue-specific- lineage, which fails to reflect the tissue complexity of eye development. Here, we describe the generation of multi-zone ocular precursor cell (mzOPC) culture derived from hiPSC in 2D, similar to other studies termed SEAM (19, 20, 23, 43, 44). In each zone of mzOPC, cells exhibited characteristics of different tissues of the developing eye: NE, SE, lens, and NC (29). But in contrast to that reported, our findings demonstrate the remarkable ability of multi-ocular organoids to self-organize from mzOPC in suspension, and to form distinct eye structures such as retina, cornea and RPE that are interconnected. Although these tissues resemble those of the developing eye, it remains to be determined whether ocular tissue could be guided to organize into its natural conformation to mimic the eye.
The aim of this study was to obtain an organoid system model able to generate complex ocular organoids from the same hiPSC culture, basically from the same individual. In our study, for the initial induction of differentiation we used a 2D culture approach to generate mzOPC for the initial stage of eye-field differentiation following previous studies on self-formed, ectodermal, autonomous, multizone (SEAM) of ocular cells from human iPS cells (19–21). The reason for initiating the differentiation from a 2D culture system instead of a 3D culture system is because 3D systems are not that effective in generating other anterior eye tissue such as cornea, although it has shown a high capacity to develop retinal organoids (15). All types of ocular progenitors are necessary to proceed with the formation of multi-ocular organoids by self-arrangement, as yet not seen altogether starting from 3D organoids. Thus, we introduced some modification to the 2D differentiation protocols based on our and other previous studies (22, 30, 45). We started with 75% confluent hiPSC cultures instead of single colonies, which led to spontaneous self-formation of NE and SE areas across the plate. The efficiency usually ranged from 60 to 120 multi-zones per well of a 6-well plate. But other SEAM protocols give rise to round multi-zones with 3–5 different concentric rings, as they were started from single colonies. In contrast, we observed that in our culture, multi-zones although usually round, also had elongated shapes that varied in size and were formed by 3–4 rings after 4 weeks of differentiation. Our typical mzOPC contains NR in the innermost ring, RPE cells in the second ring with a region of NC between NR and RPE, and SE in the last ring, sometimes containing lens cell clusters, similar to SEAM organization (19, 21). During differentiation, cells constantly interact with other cells, the extracellular matrix and other components. Interactions can be mediated through direct cell-to-cell, cell-to-matrix contacts or secreted factors. Thus, differences in the cellular differentiation, maturation and migration of our mzOPC compared with other SEAM might be attributable to the extracellular matrix coating (43). In our system, the spontaneous mzOPC organization seemed not to be conditioned by the extracellular matrix coating, as we used Matrigel as well as Geltrex as a substrate for mzOPC differentiation with similar results (not shown).
Following the formation of individual ocular tissue-specific areas within the mzOPC stage, the subsequent stages of ocular organoid development in 3D were driven by self-refolding of ocular structures preventing its disruption and maintaining cell-cell interactions, cell number and cell signaling. Using this system, the organization of the different ocular tissues within the organoids were random and did not recreate the primitive eye conformation. This is probably due to the starting point of the differentiation in 2D and the technique to lift up 2D structures.
During early eye development, retinal progenitors emerge from PAX6 + cells in the early eye field, which in turn require RAX and CHX10 to proliferate (46). Likewise, retinal cells differentiated as 2D monolayers in the mzOPC expressed important retinal markers (PAX6, RAX, LHX2, SIX3, and SIX6), and the retinal neuroepithelium self-organized to form NR structures (47). We observed that these NR regions were composed of a heterogeneous mix of retinal cells containing each major retinal cell type that organized into laminated structures. Once lifted, the NR formed retinal organoids consisting of layered structures with the main photoreceptor subtypes including photoreceptors with Rhodopsin, L/M-opsin and S-opsin in the outer retina, and ganglion cells in the inner part emulating many of the temporal and spatial characteristics of in vivo development (48). During human retinal development, photoreceptor OS emerge and elongate at weeks 16–20 of gestation coinciding with the emergence of OS in our retinal organoids observed at day 140, although the OS observed had very few or disorganized disk stacks likely due to the immaturity of the organoid. Longer incubation periods, up to 200 days, is described to enhance photoreceptor OS maturation (49). However, our results show that our retinal organoids can develop some synaptic maturity at day 140. Electron microscopy data showed the presence of electron-dense ribbons surrounded by synaptic vesicles, and Synaptophysin and vGlut1 expression, confirming synapses between retinal layers (50, 51).
Corneal organoids, developed from SE, become key 3D models to understand the corneal development process and diseases (52). The emergence of SE during the differentiation of hiPSC into eye-field commitment is related to signals that regulate eye development. The transcription factor p63, especially its isoform ΔNp63α, has been linked to stemness and is highly expressed in the basal layers of the corneal epithelium and in the limbus (53). The presence of p63 + cells together with PAX6 in our mzOPC and organoids suggests a primitive epithelial surface (44) similar to those obtained from human limbal cells (54). It is described that inhibition of BMP4 during SEAM differentiation decreases expression of p63 (44) but, in contrast, the presence of noggin (inhibitor of BMP4) during the first 30 differentiation days in our mzOPC cultures did not block p63 expression or the formation of surface ectoderm. Upon further differentiation, SE areas formed corneal-like organoids with a transparent epithelium filled with fluid and subsequently stroma, and expressed proteins specific to cornea, conjunctiva and limbus (15, 23). Given the common developmental origin and a degree of structural similarity, both cornea and conjunctiva are transparent multilayer epithelia with some distinct features, such as cell layer thickness, type of cells, or the presence of mucins for the tear film (55), but it was not possible to distinguish by eye which ones were corneal or conjunctival. To further verify the maturation of corneal cells in our organoids, we confirmed the protein expression of CK3, AQP1 and N-Cad to distinguish between corneal and conjunctival epithelia (35, 56). We did observe a certain degree of stratification after 140 days, supporting the conclusion that these cells are able to mature toward terminally differentiated and stratifying corneal epithelial cells. Although we observed some corneal regions, our organoids contained mostly conjunctival epithelium expressing CK5, CK19, Na+/K+-ATPase and p63, which have been previously established as putative markers (57). Moreover, conjunctival-predominant corneal organoids contained stratified columnar-type cells with interspersed goblet-like cells expressing mucin 1. Corneal organoids also contained mesenchymal cells in addition to ocular-specific cells (15, 23). Mesenchymal cells might be derived from NC cells, rather than from mesoderm cells, that appeared during differentiation. It is likely that NC-like cells come from proximal cells to the ectoderm, such as periocular NC, during eye development (5), which might be important for the development of organoids and epithelial growth. Indeed, we found that VIM + keratocyte-like cells produce stromal collagen types I and V, important to shape corneal organoids, generating a subepithelial basement membrane-like structure that resembles the Bowman layer.
Previously, other studies generated RPE spheroids that differ from our RPE organoids (58–62). They obtained compact RPE spheroids from ARPE19 or primary RPE cell aggregation that did not acquire apical-basolateral polarity, nor form a Bruch’s membrane-line, collagen-filled or fluid-filled stroma, which did not recapitulate the native RPE epithelium. In contrast, we and Liu et al., (41) generated RPE organoids consisting in a RPE monolayer on the surface, with microvilli in the apical side and deposits of collagen in the basolateral side that resemble the Bruch’s membrane. RPE organoids acquired pigmentation over time and could be isolated and purified to obtain an RPE cell monolayer, which can be easily amplified and passaged retaining a proper RPE phenotype and function, such as phagocytosis of photoreceptor OS, production of VEGF and epithelium barrier (41). This makes them useful for cell therapy either by single cell injections (30, 31) or by transplantation a part of the RPE organoid cut in two as they are already polarized and can be integrated, similar to RPE in sheets or scaffolds (63), but using its own ECM as a support.
Several creative approaches are being developed to generate more advanced organoids. Some strategies that recently appeared consist in the fusion of organoids that have been grown apart, replacing the undefined Matrigel matrix with defined synthetic hydrogels and guided self-organization (64). Organoids generated using our self-organization protocol display a high complexity, containing different eye parts -retina, RPE and cornea- that could derive from the ectoderm germ layer. One further step to achieve even more complex and complete organoids would be to incorporate blood vessels (65) and microglia (66), arising from the mesoderm. These components would be crucial as multi-lineage communication is required for tissue development to allow the different cell types to remain in contact and, therefore, contributing to tissue maturation and further differentiation of the organoid (67). Thus, the current key limitation of our protocol is the lack of integrated vasculature or immune cells, necessary to fully mimic adult ocular tissue composition (68). The presence of these extra cell types in the stage of our multi-ocular organoids is not relevant to emulate early ocular development. Despite that, for later stages of development these components would be crucial, as multi-lineage communication is required for tissue development contributing to tissue maturation (69). To achieve this issue, our differentiation protocol should induce ectodermal and mesodermal lineages simultaneously, to be able to obtain neovascular niche or primitive hematopoiesis (66), considering that they do not arise under the same differentiation conditions. The addition of vasculature (67, 69) or microglia (70) (either by single cell addition or by organoid assembly (71)) to integrate within the ocular organoids while they are forming will overcome the aforementioned limitation.
Blindness can be caused by many factors, from genetic defects, injuries of the anterior segment of the eye such as corneal scars or infections, conjunctivitis, cataract or keratitis, to posterior segment diseases such as age-related macular degeneration, diabetic retinopathy or glaucoma. The different ocular organoids developed in this study displays a potential use in developmental biology studies, disease modeling, drug testing and personalized medicine.
{kind=link}