Hulless barley (Hordeum vuglare L. var. nudum Hook. f.), also called naked barley, is an important cereal crop in Tibet Plateau(Xu et al., 2016). It has been served as a healthy food for human consumption and animal feed for over thousands of years. Owing to high altitudes, the naked barley is cultivated in harsh environment such as valleys and higher land on Tibet(Liang et al., 2017). It is also affected by drought and low temperature in March every year when the weather is quite cold and dry(J. B. Du et al., 2011). In order to mitigate adversities such as drought, salinity, and low temperature, various strategies have been evolved in hulless barley(H. Li, Guo, Lan, Zhou, & Wei, 2014).
It is well-known that drought is the most serious environmental stress that affect crop growth and yield by causing a wide range of physiological and biochemical responses(Flowers, 1989; Iqbal, Murtaza, Saqib, & Ahmad, 2015). The influences of drought on the crop varies between varieties and developmental stages. Many genes responsible for drought tolerance have been reported for decades(Lenka, Katiyar, Chinnusamy, & Bansal, 2015; Moa, Liua, Lina, Xub, & Xianga, 2005). For example, the transcription factor AtHB13 has been reported to act as a positive regulator of drought tolerance in Arabidopsis. Genetically, drought tolerance is extremely complex trait involving in several genetic pathways such as polygenic control and complex morpho-physiological mechanisms(Xuekun Zhang et al., 2014). At the molecular level, drought stress could induce genome-wide changes in gene expression via epigenetic mechanisms like histone modification and DNA methylation in plants(Wang et al., 2016; Zong, Zhong, You, & Xiong, 2013). For example, the mutant allele (met1) DNA methyltransferase 1 locus could remove methylation at several genomic regions leading to specific expression of 31 stress response-related genes in tobacco(Ku, Lim, & Park, 2006; Zubko, Gentry, Kunova, & Meyer, 2012).
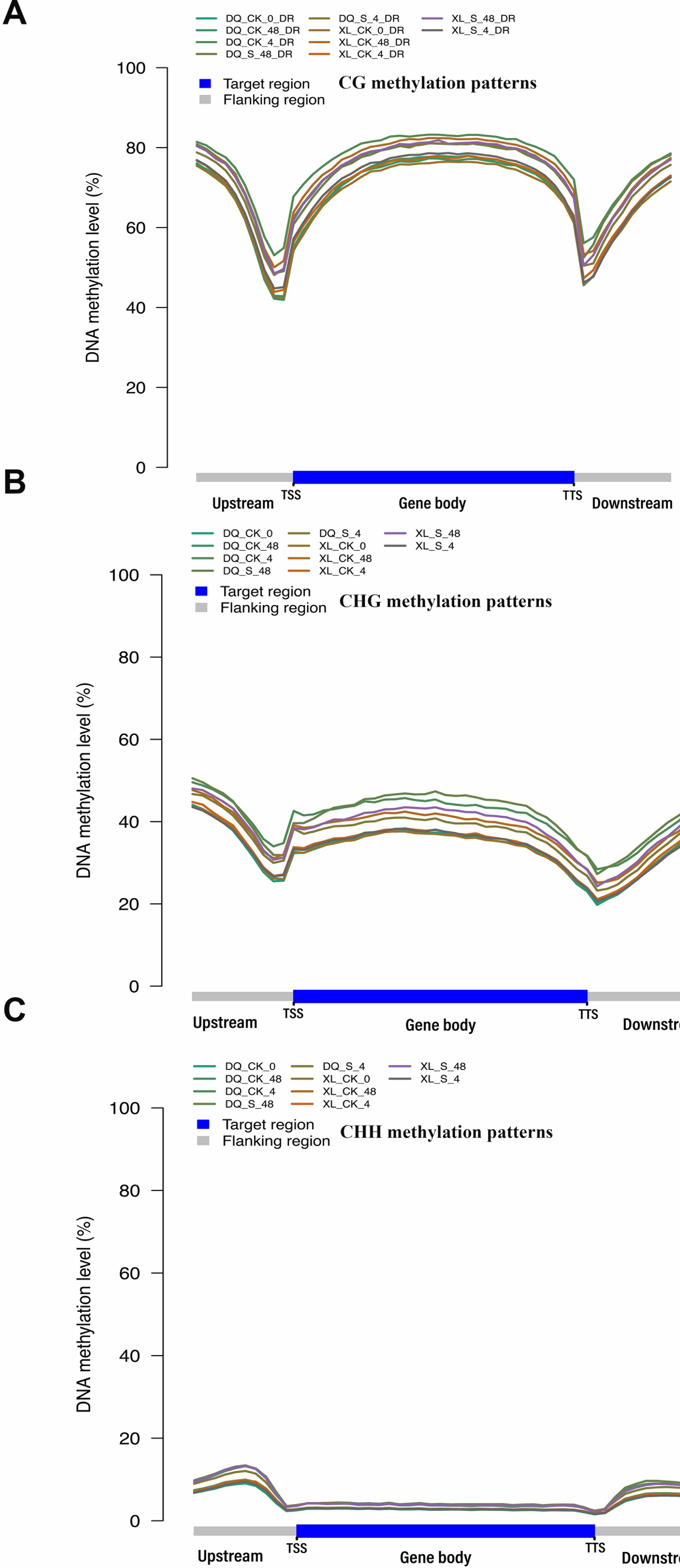
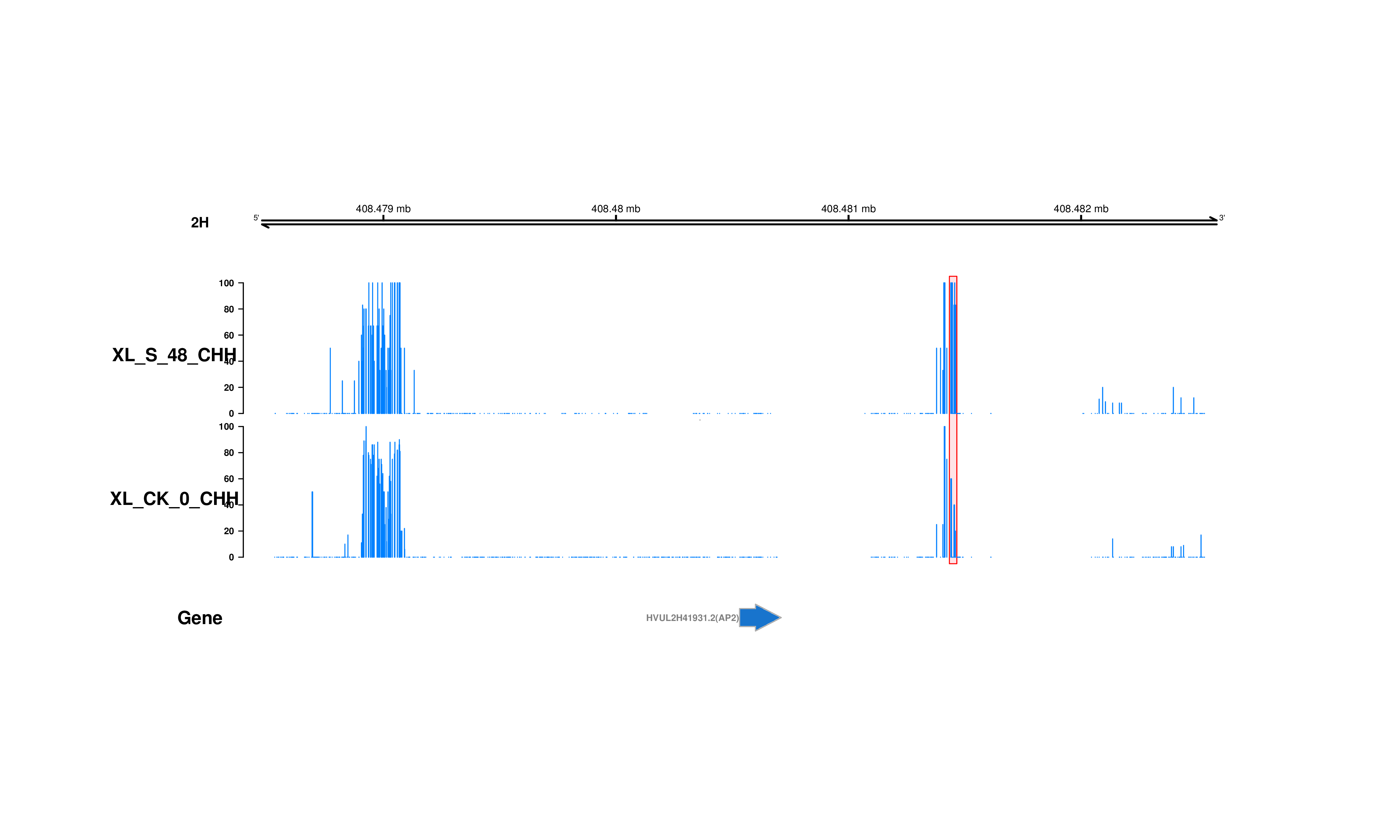
DNA methylation, an epigenetic modification, play crucial roles in plant growth and development as well as responses to various abiotic stresses(Arthur et al., 2018; Uthup, Ravindran, Bini, & Thakurdas, 2011). Actually, DNA methylation exists in all eukaryotes, and it frequently occurs at the 5-position of cytosine (5mC)(Kasai & Kawai, 2009). Under normal conditions, the proportion of methylcytosine in plants is 20–30% in plants, which usually occurs in three nucleotide sequences: CG, CHG and CHH (H indicates C, T or A)(Serre, Lee, & Ting, 2009). CG methylation is produced by the conserved DNA methyltransferase METHYLTRANSFERASE1 (MET1)(Zubko et al., 2012); CHG methylation is modified by the plant specific DNA methyltransferase CHROMOMETHYLASE3 (CMT3)(J. Du et al., 2012); and the de novo CHH is methylated by 24-nucleotide small interfering RNA dependent DNA methylation (RdDM) pathway(Ma et al., 2015). Interestingly, methylation levels contribute greatly to the process of adaptation to stress in plants. The hyper or hypo methylation changes in the hybrids could be an indicator of the expression levels of stress related gene under drought, when compared to their parents(Boyko et al., 2010; Hai & Zhang, 2009). This indicates the methylcytosine and its reversibility may regulate transgenerational response to stresses. An increasing number of studies revealed the transposon-rich heterochromatic regions show heavily methylation(Melamed-Bessudo & Levy, 2012). Over one-third of expressed genes are methylated within transcribed regions, while only 5% genes within their promoter regions in Arabidopsis(Xiaoyu Zhang et al., 2006). Thus, DNA methylation within genes is a common feature of eukaryotic genomes. Intriguingly, methylation of transcribed regions does not usually result in gene silencing(Xiaoyu Zhang et al., 2006). It seems to primarily occur at CG sites and appears to show moderate correlation between the level of gene-body methylation and gene expression(Su, Wang, Xing, Liu, & Yong, 2014). DNA methylation shows the deposition of certain chromatin marks such as differentially modified histones because of the tight link between DNA methylation and histone modifications(Fuks, 2005). For instance, there is a relationship between the reduce in DNA methylation levels and the changes in level of H3K4me and H3K9me modifications in DECREASE IN DNA METHYLATION1 (DDM1) mutant, which maintain cytosine DNA methylation within the heterochromatic locus regions(Sasaki, Kobayashi, Saze, & Kakutani, 2012; Zhou et al., 2013).
It is the first time that the epigenetic responses to drought between two contrasting varieties were revealed via bisulfite sequencing and RNA sequencing in Tibetan hulless barely. Here, we explored how DNA methylation is involved in drought responsiveness. Changes in the level of DNA methylation affect plant resistance to drought, especially the methylation changes of several transcription factor genes. Therefore, epigenetic changes in genome could be considered as an important regulatory mechanism for plants to adapt to drought and possibly other environmental stresses.
{kind=link}
{kind=link}