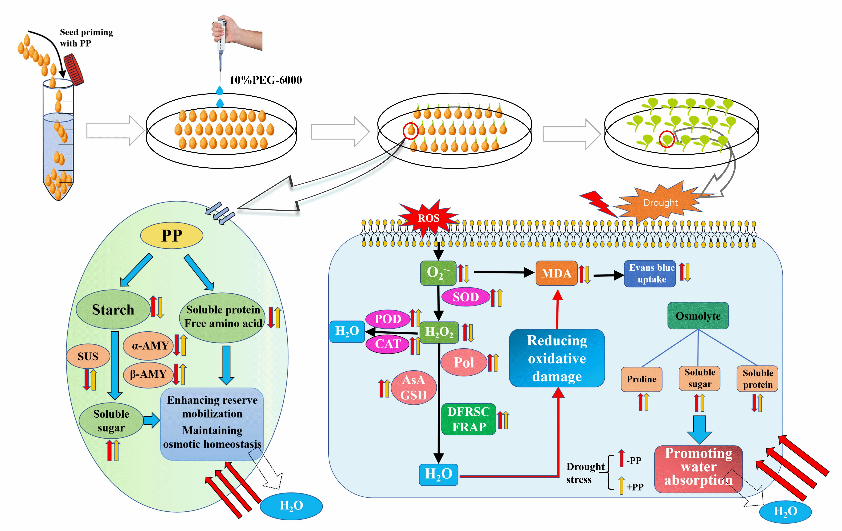
Drought is a main abiotic stress that threatens plant growth, development, and yield production by inhibiting various physiological processes (Fabregas and Fernie, 2019; Razi and Muneer, 2021; Trenberth et al., 2013). Seed germination and seedling growth, as the critical stages of plant growth, are susceptible to drought stress, usually exhibiting low germination rates, inhibition of root elongation, and biomass reduction (Marthandan et al., 2020; Finch-Savage and Leubner-Metzger, 2006). In the present study, drought stress induced by the usage of 10% PEG significantly reduced the germination rate, germination potential, and germination index of tomato seeds, as well as the seedling fresh weight, seedling height, and root length (Fig. 1). In order to reduce the negative effects of drought stress on seed germination and seedling growth, a low-cost and feasible method known as seed priming has been widely used to improve drought stress tolerance (Marthandan et al., 2020; Salah Sheteiwy et al., 2018; Zulfiqar, 2021). Remarkably, seed priming with PP significantly alleviated drought-induced inhibition of seed germination and seedling growth. Notably, this positive effect of PP application is perhaps due to the high supply of active peptides and amino acids, which confer crop tolerance to various abiotic stresses by regulating biochemical and physiological processes (Colla et al., 2015). Therefore, to the best of our knowledge, our study is the first to verify that PP could alleviate the reduced tomato seed germination and seedling growth; thus, it could be a potential strategy to improve seed tolerance to drought stress.
Seed germination is a complex physical and chemical process, beginning with water uptake of dry seeds to radicle protrusion and growth, where the necessary energy is supplied by the degradation of storage substances, such as starch and protein (Lei et al., 2021; Finch-Savage and Leubner-Metzger, 2006). It has been widely indicated that seed reserve content is positively correlated with the germination rate and seedling growth of wheat (Triticum aestivum L.), rice (Oryza sativa L.), and lentil (Lens culinaris Medikus) (Biju et al., 2017; Cheng et al., 2015; Lei et al., 2021). However, mobilization of starch and protein is prevented under abiotic stresses, including drought (Fabregas and Fernie, 2019; Marthandan et al., 2020). The present study showed that drought stress remarkedly inhibited the activities of α-amylase, β-amylase, total amylase, and sucrose synthase, leading to a reduction in the hydrolysis of starch to soluble sugars (Fig. 2 and Fig. 3). In addition, the PEG-induced water deficiency also adversely decreased the content of soluble protein and free amino acids, suggesting that drought stress inhibited the metabolism of sugar and degradation of storage proteins during germination. However, PP priming significantly increased the activities of amylase and sucrose synthase and the levels of soluble sugar, soluble protein, and free amino acids in tomato seeds. These results indicate that PP may enhance amylase activity and subsequent reserve mobilization to stimulate seed germination under drought stress. Previous findings have revealed that protein hydrolysates are closely associated with the levels of endogenous hormones, as their biosynthesis, stimulating the activity of various enzymes, including amylase (Casadesús et al., 2020; Sorrentino et al., 2021).
Plants usually accumulate a variety of substances for osmotic regulation, such as proline, betaine, soluble protein, and soluble sugar, which help to maintain turgor pressure, promote water absorption and retention, and improve tolerance among plants against drought stress (Ozturk et al., 2021; Razi and Muneer, 2021). In the present study, rapid increases in soluble sugars, soluble protein, and proline accumulation in tomato seedlings were also observed under drought conditions. Seed priming with PP resulted in further improvements in soluble sugars, soluble protein, and proline accumulation (Fig. 3 and Fig. 7), which may be beneficial for the osmotic homeostasis under drought stress. Similar to our results, exogenous application of silicon, quercetin, and melatonin partially improved drought resistance via increasing the levels of soluble sugars, soluble protein, and proline in plants (Lei et al., 2021; Patel et al., 2021; Yang et al., 2021). Proline, an amino acid act as an essential osmolyte, and its accumulation in drought-treated seedlings after PP priming might be due to the high proline supply of PP, which contained 2.45% of proline (Fig. S1). Therefore, it can be speculated that PP priming might play a key role in seed tolerance to drought stress by improving the accumulation of active osmolytes, thereby reducing osmotic toxicity.
Under normal growth conditions, the production and removal of ROS are in a dynamic balance, which does not cause damage to plants in general (Choudhury et al., 2017). However, plant standard homeostasis is remarkably perturbed because of the drought-caused excessive generation of ROS, such as O2•−, 1O2, and H2O2 (Marthandan et al., 2020). The results of this study showed that the contents of O2•− and H2O2 significantly increased under drought stress, causing an excessive production of ROS (Fig. 4a and b). Overproduction of ROS under abiotic stress usually leads to an oxidative damage to the cellular components, leading to membrane lipid peroxidation and cell membrane destruction (Kamal et al., 2021; Sasi et al., 2021; Sun et al., 2017). The MDA and Evans blue uptake are considered as key indicators of lipid peroxidation and integrity of the plasma membrane in plants to reflect the degree of membrane damage (Sun et al., 2017). Under drought stress, the augmented MDA and Evans blue uptake were observed compared to the control (Fig. 4c and d). However, PP priming significantly reduced the contents of O2•− and H2O2 in seedlings, thereby obviously minimizing the oxidative damage, as indicated by the reduced MDA and Evans blue uptake under drought stress (Fig. 4a-d). Moreover, histochemical staining also clearly demonstrated that drought stress could cause rapid accumulation of O2•− and H2O2, resulting in a subsequent oxidative damage to the cell membrane in the root tips; however, PP priming could effectively reduce the widespread staining of root tips (Fig. 4e-g). Altogether, these data indicate that seed priming with PP enables tomato seedlings to maintain ROS at an appropriate level, thereby contributing to enhanced drought tolerance.
To overcome oxidative damage, plants invoke efficient antioxidant defense mechanisms including enzymatic and non-enzymatic antioxidants (Fang and Xiong, 2015; Khan et al., 2021). As the first line of defense mechanisms, SOD is an important enzyme in catalyzing the disproportionation of O2•− to H2O2. Furthermore, the H2O2 produced in response to superoxide dismutase or other metabolic activities can be reduced to H2O by the action of CAT and POD, thereby decreasing ROS toxicity (Fang and Xiong, 2015; Patel and Paroda, 2021). It was noticed that the activities of SOD, POD, and CAT in tomato seedlings were largely upregulated after the PP priming, compared to the PEG treatment alone (Fig. 5a). The results in this study are in accordance with Sitohy et al. (2020), who indicated that pumpkin seed protein hydrolysate treatment could enhance CAT and SOD activities, which is positively related to salt tolerance. Moreover, other ROS scavenging antioxidants are a class of low molecular weight compounds such as phenolic compounds, carotenoids, GSH, and AsA, which can effectively scavenge the accumulation of ROS in plants, and thus protect them from oxidative damage under abiotic stresses (Li et al., 2021; Zhou, et al., 2018). In our study, drought stress substantially increased the contents of total phenolics, flavonoids, anthocyanins, AsA, and GSH in tomato seedlings, whereas PP priming further improved these antioxidant contents (Fig. 5b). Consequently, this results in a considerable increase in antioxidant activities as indicated by DFRSC and FRAP in tomato seedlings (Fig. 6). In addition, a large number of antioxidant peptides have been identified from different sources of protein hydrolysates that exhibit strong antioxidant capacity (Bougatef et al., 2010; Wen et al., 2020). Therefore, the decreased ROS accumulation in tomato seedlings after PP priming might be partially due to the presence of some antioxidant peptides in PP. Subsequently, these results indicate that seed priming with PP effectively scavenge ROS through improving the activities of antioxidant enzymes and accumulation of antioxidant compounds, and consequently enhancing the tomato seed tolerance to drought stress.
{kind=link}