Plant pathogens can have devastating effects on a wide range of food crops and are responsible for a number of pandemics causing catastrophic effects on social stability such as the Irish and Bengal famines [1]. In addition, mycotoxins synthesized by some pathogens such as Aspergillus flavus and Fusarium verticillioides can directly harm human and animal health [2]. Therefore, the effective management of plant pathogens is a paramount task for ensuring human health and social stability. The introduction of synthetic fungicides revolutionizes agricultural production both in quantity and quality by providing highly effective management of plant diseases [3, 4]. However, repeated and excessive application of same active compounds over large spatial scales can lead to the development of fungicide resistance in pathogen populations, rapidly rendering efficacy to manage plant diseases [5, 6].
Synthetic fungicides can be classified to site-specific and site non-specific (multisite) according to modes of action. Site-specific fungicides are highly active and often systemic (taken up by and distributed throughout plants), resulting in good disease mitigation at very low dose to specific fungal groups. Site non-specific fungicides, usually having a broader breadth of metabolic activity, can be used to prevent or eradicate a wider range of plant pathogens. Resistance to site-specific fungicides can occur as a result of changes in a single amino acid of the target protein in the pathogen, whereas for site non-specific fungicides the development of resistance involves multiple nucleotide changes across several targeted genes in the pathogen genome. Therefore, it is commonly believed that the risk of developing resistance to site non-specific fungicides is lower than that to site-specific fungicides [3]. However, mutations to fungicide resistance may generate fitness penalties that reduce the competitiveness of pathogens in the absence of the corresponding fungicide(s) as a result of the changes in DNA sequences impeding their important cellular and biochemical functions [7, 8]. Thus, appropriate implementation of fungicide in agricultural practices that minimizes directional selection on pathogen populations by mixing or rotating fungicides with different modes of action is thought to an effective approach to slowdown the development of field resistance.
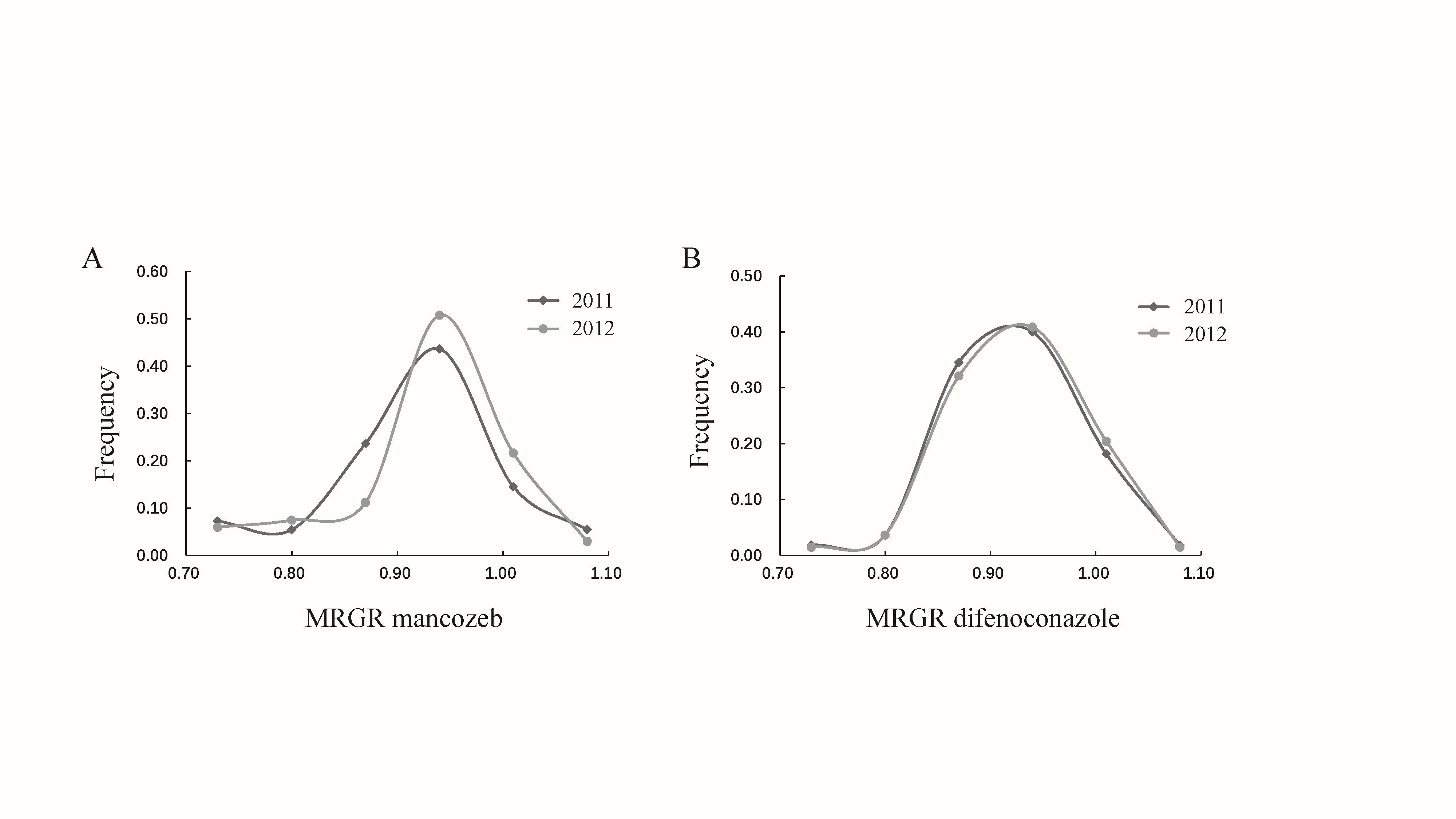
Pathogen populations that develop resistance to one fungicide sometimes can simultaneously resist one or several other fungicides-a phenomenon known as cross-resistance. Usually, cross-resistance appears among fungicides with the same mode of action [9, 10]. For example, Alternaria alternata the causal agent of leaf blight of pistachio was reported to show cross-resistance to difenoconazole, propiconazole, and tebuconazole, all of these fungicides act through demethylation inhibition (DMI) [11]. However, cross-resistance may also occur between fungicides with distinct modes of action [12]. This is usually caused by limiting intercellular accumulation of active compounds through enhanced drug efflux and detoxification or reduced drug uptake. For example, the overexpression of efflux transporter genes made the grey mould fungus Botrytis cinerea simultaneously resistant to a broad-spectrum of fungicides [13]. In field isolates of Zymoseptoria tritici, enhanced efflux contributed to the pathogen’s cross-resistance to several fungicides with different modes of action [14]. In this study, we used efficacy profile of mancozeb and difenoconazole in Alternaria alternata from potato to test the development of cross-resistance to fungicides with different modes of action.
Mancozeb is a dithiocarbamate fungicide classified by the Fungicide Resistance Action Committee (FRAC) to mode-of-action group M (Multi-site Action). Mancozeb itself is not fungicidal, but the ethylene bisisothiocyanate sulfide (EBIS) and ethylene bisisothiocyanate (EBI) generated after its exposure to water and UV light are active toxicants which interfere with sulphydryl groups of enzymes involving at least six biochemical processes within cytoplasm and mitochondria of fungal cells [15]. On the other hand, difenoconazole, a 1, 2, 4-triazole, is a demethylation inhibitor (DMI) that target sterol 14𝛼-demethylase (CYP51), an important regulatory enzyme in the ergosterol biosynthetic pathway [16].
Mancozeb and difenoconazole are routinely used together to control plant diseases in many parts of the world. As a result, many studies have dedicated to understand their efficacy of mitigating disease epidemics and molecular mechanisms contributing to the development of mancozeb and difenoconazole resistance in pathogens [17, 18]. Although the risk of developing mancozeb resistance is low, substantial reduction in sensitivity to the fungicide has been documented in many pathogen species [15]. Genome-wide analysis reveals that this reduction in pathogen sensitivity is associated with the genes involving the formation of transcriptional machinery, cellular pH regulation, and multidrug transporters [17]. For difenoconazole, resistant phenotypes have been reported in many field populations of fungal pathogens [19, 20]. Up-regulation of ABC or MFS transporters to increase efflux, alterations of the Cyp51 gene to decrease the affinity of DMIs for their target site and the raised levels of sterol 14α-demethylase caused by overexpression of the Cyp51 gene are the three main mechanisms responsible for the final phenotype of difenoconazole resistance in plant pathogens [13]. However, information concerning the evolutionary interaction of resistance to the two fungicides in pathogens and how such interaction may impact the development of pathogen’s aggressiveness is limited but important to effectively administer fungicides for sustainable food production and social development.
Mancozeb and difenoconazole are also commonly used together to control potato early blight, a foliar disease forming dark-brown to black necrotic lesions with concentric rings [21]. The disease occurs worldwide but is most prevalent and severe in areas experiencing warm and alternating dry and high humidity periods [22]. Potato early blight can be caused by several species of Alternaria [23], but A. alternata, a smaller-spore species which produces conidia containing 8-12 spores with numerous secondary and occasionally tertiary chains branching from apical and median cells [24], is the main causal agent in China [25, 26]. No teleomorphs (sexual fruiting body) have yet been detected either in the field or under laboratory conditions, but population analyses of genetic variation, mating type distribution and phylogenetic trees all suggest that sexual reproduction or some other mechanism beyond mutation must occur to generate genetic variation of the pathogen for ecological adaptations [25]. High genetic variation in the pathogen also increases its potential to develop fungicide resistance, posing a great threat to the long-term management of the pathogen.
Thus, the specific objectives of this study were to: i) determine the spatial distribution of A. alternata sensitive to mancozeb and difenoconazole in China; ii) assess cross-resistance of mancozeb and difenoconazole sensitivity in A. alternata; and iii) evaluate whether there is a fitness cost associated with fungicide resistance by conducting correlation analyses of mancozeb and difenoconazole tolerances with pathogenicity in A. alternata.
{kind=link}