Despite significant improvements in diagnosing hepatocellular cancer through various methods, appropriate novel strategies, such as specific molecular regulatory mechanisms, are still required to optimize the treatments and diagnosis of HCC33. With the advent of the bioinformatics era and increased research on cancer-related genes, it has been demonstrated that there is a network of complex interactions between RNAs known as the ceRNA network (ceRNET) in the way that is focusing on this network and the factors involved, may promote the rapid diagnosis of various malignancies34.
Thousands of highly-conserved sequence circRNAs have been identified thanks to advances in high-throughput sequencing technology and computational biology techniques. These circRNAs, as a potential regulatory genes, may act as ceRNAs in cell physiology, trapping miRNAs and thereby influencing the regulation of other transcripts35. Various studies have shown that circRNAs are expressed differentially in tumor tissues, including circRASGRF236, CircPUM137, and circGFRA138, which have been related to HCC progression.
Similarly, there may currently be many circRNAs in HCC that need to be studied, identified, and assessed. CircRNAs, on the other hand, have excellent tolerance to exoribonuclease due to their covalently closed-loop structure. These molecules may be used as efficient diagnostic and therapeutic targets and as promising biomarkers for prognosis39.
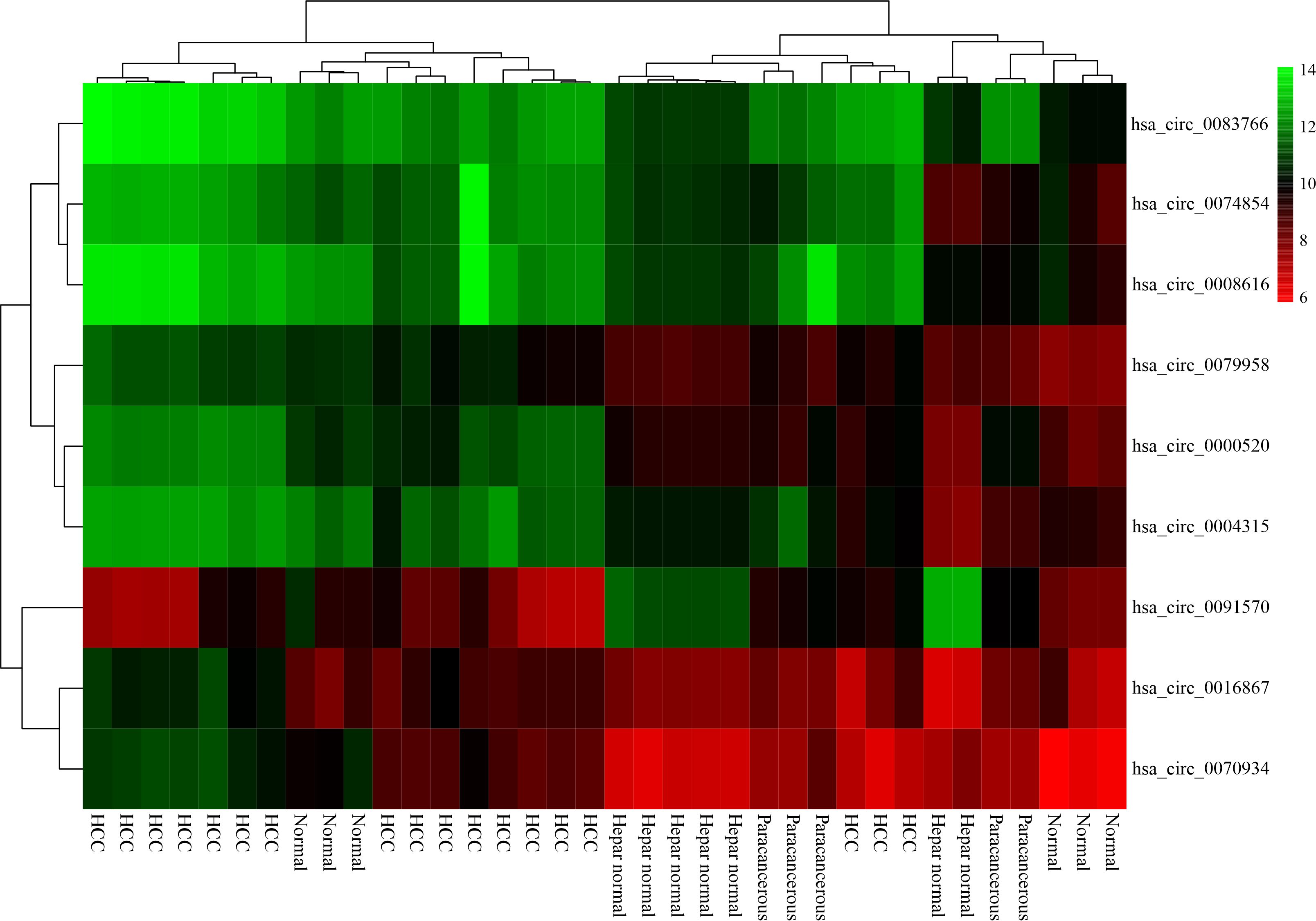
In this study, we first gathered three circRNA microarray datasets from the GEO database, and then we screened nine DECs in the initial stages. (Table.3) Other researchers have investigated the role of hsa_circ_0074854 and hsa_circ_0091570 in hepatocellular cancer40,41. However, the other seven DECs in liver cancer have yet to be investigated.
These seven DECs were found to be abnormally upregulated in HCC cells based on our findings. According to the ceRNA hypothesis, increased expression of these seven DECs may decrease the expression of miRNAs inside the cell. We predicted DEC-related MREs by using intersecting, miRNAs from the CSCD database and downregulated DEMIs. The results revealed that only four DECs (hsa_circ_0000520, hsa_circ_0004315, hsa_circ_0008616, hsa_circ_0070934) were ascertained as ceRNAs to downregulate miRNA expression inside the cell. These miRNAs, includes hsa-miR-542-5p, hsa-miR-326, hsa-miR-511-5p, hsa-miR-195-5p, hsa-miR-214-3p, and hsa-miR-424-5p were named FImiRNA in this research.
Among these six FImiRNAs, miR-542-5p has a complementary sequence with MRE sites related to two DECs, hsa_circ_0000520 and hsa_circ_0008616. According to the previous research, miR-542-5p expression is downregulated in non-small cell lung cancer (NSCLC) tissues, which is implicated in NSCLC tumorigenesis42. This miRNA has also been investigated in breast cancer, endometrial carcinosarcoma, and osteosarcoma43–45.
For miR-326, some studies have found that this miRNA acts as a tumor suppressor in glioblastoma, human prostatic carcinoma, and breast cancer. The expression of miR-326 is downregulated in these cancers46–48. Besides that, this miRNA has been demonstrated to reverse chemoresistance in human lung cancer by targeting the specificity protein 1(SP1)49.
MiR-511-5p has been studied for its tumor-suppressive role in preventing colorectal cancer cell progression by targeting GPR11650 and lung adenoma-carcinoma cells by targeting oncogene TRIB251. Also, a study has proven that miRNA-511-5p prevents malignant behaviors of breast cancer with a direct effect on SOX9 and PI3/AKT regulatory pathway52.
For miR-195-5p, overexpression of this miRNA has been demonstrated to inhibit cell migration and invasiveness in oral squamous cell cancer, cervical cancer, and breast cancer by targeting TRIM14, YAP1, and CCNE1 proteins, respectively53–55, or preventing thyroid carcinoma cell progression by acting on the p21/cyclin D1 axis56. Overall, miR-195-5p has the potential to be a tumor suppressor. MiR-214-3p and miR-424-5p, like the previously mentioned miRNAs, were investigated in some research for their involvement in reducing cell progression through their effect on transcripts and signaling pathways. MiR-214-3p, for example, serves as a tumor suppressor by targeting ABCB1 and XIAP proteins, preventing multi-drug resistance and stimulating apoptosis. MiR-424-5p as well, suppresses intrahepatic cholangiocarcinoma metastasis by targeting ARK557–61. In summary, the outcomes of this study on these miRNAs are similar to other studies, and these miRNAs may play a critical part in the development of HCC.
To determine the effects of DECs on mRNAs through miRNAs, an intersection was performed between the overlapped target genes of six FImiRNAs from the Mirwalk and Targetscan databases and upregulated DEGs from the TCGA; as a result, 543 genes were obtained and named as FImRNAs. Afterward, a circRNA/miRNA/mRNA regulation network was established as a ceRNA network based on DEC-FImiRNA and FImiRNA‐FImRNA interactions.
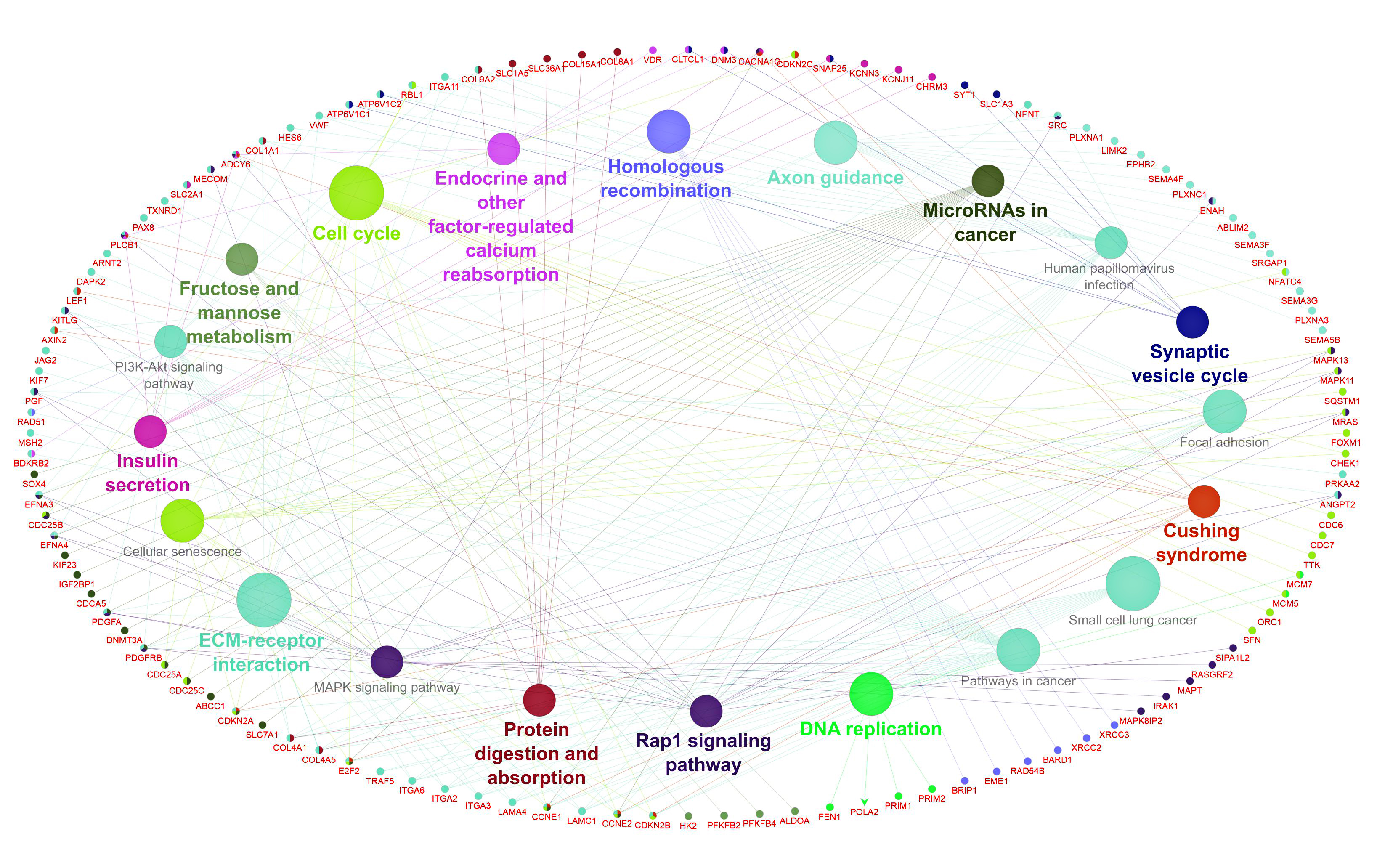
The GO and KEGG databases were employed to understand the biological roles and potential function of 543 FImRNAs. Based on GO analysis, these genes in the biological process are mainly enriched in the positive regulation of the cell cycle, DNA replication, and nuclear division. Therefore, they may play pivotal roles in tumorigenesis and tumor development. While KEGG analysis showed that these genes were mainly enriched in the ECM-receptor interaction, cell cycle, and Rap1 signaling pathway. these pathways have been examined in some studies that have shown many roles of these genes in cancer progression. In conclusion, the circRNAs explored in this study may perform similar or related functions via the circRNA-miRNA-mRNA axis.
Rap1 has a variety of functions in tumor initiation and development. Rap1 regulates integrins and cadherins by stimulating EGFR and Src/FAK, which plays essential roles in cell adhesion to ECM and cell-cell adhesion; both are crucial for tumor cell invasion and metastasis. Also, Rap1 induces tumors and epithelial-mesenchymal transition (EMT) via notch signaling. During tumorigenesis, the interaction between cancer cells and the tumor microenvironment (TME) causes ECM stiffness and alteration of ECM key receptors, leading to aberrant mechanotransduction and malignant transformation.
On the other hand, Src triggered by Rap1 signaling may activate the MAPK/ERK pathway, which has been found in some research to promote G0/G1 to S phase cell cycle progression and angiogenesis in cancer62–65. Thus, these FImRNAs' function and signaling pathways are associated with the occurrence and development of tumors. In summary, these FImRNAs, which are indirectly regulated by the four selected DECs we identified, may play a vital role in the HCC signaling pathway.
When 543 FImRNAs and 441 survival-related DEGs were intersected, only 204 FImRNAs were determined to be significant in survival. We built a PPI network for these 204 genes and selected ten hub genes from the network for further analysis. Fig. 10A shows the overall survival time of ten hub genes in Kaplan-Meier plots. A circRNA-miRNA-mRNA regulatory axis was created for these hub genes to help the researchers understand them better. The top five hub genes based on the most significant p-value among these ten hub genes that considerably affect overall survival are KIF20A, NCAPG, TTK, PLK4, and CDC6.
KIF20A mRNA, which encodes protein MKlp2, has been reported to be significantly expressed in large human hepatocellular cells. MKlp2 accumulation is linked to abnormal hepatocyte proliferation and tumor aggressiveness in human hepatocellular carcinoma66. Interestingly, several studies showed a correlation between KIF20A overexpression and tumor progression and proliferation in colorectal cancer, renal clear cell carcinoma, and bladder cancer cells66–68.
NCAPG expression is increased in various cancers, including hepatocellular liver cancer, as an oncogene that stimulates cell proliferation and apoptosis through the PI3K/AKT/FOXO4 pathway. Another study found that knocking down NCAPG as a mitotic gene may prevent HCC cell growth, progression, and migration69,70.
According to the results of the previous studies, TTK activates Akt/mTOR, and MDM2/p53 in a p53-dependent mechanism increases cell proliferation and migration, which stimulates malignancy71. TTK overexpression has also been shown to increase HCC cell drug resistance to sorafenib, suggesting it may be a viable therapeutic target for human hepatocellular carcinoma72.
Concerning PLK4 and CDC6, their association with HCC has been described in previous researches; for example, PLK4 overexpression promotes cell growth, while its knockdown suppresses progression, invasion, and migration. CDC6 dysregulation has a role in developing many cancers, like hepatocellular carcinoma. CDC6, which acts as a regulator in the early stages of DNA replication and as a checkpoint controller before mitosis, has been associated with the clinical progression of HCC and may be utilized as a biomarker in patients with this kind of cancer73,74.
Finally, a circRNA-miRNA-mRNA regulatory axis based on DEC-FImiRNA and FImiRNA-hub gene interactions was built. This network, which contains two DECs (hsa_circ_0070934 and hsa_circ_0004315) and three FImiRNAs (hsa-miR-511-5p, hsa-miR-214-3p and hsa-miR-195-5p), along with five final hub genes, may play an essential role in the development and progression of HCC (Fig .10B). Our results suggest that these two DECs identified by bioinformatics techniques may be utilized as effective diagnostic and valuable prognostic biomarkers.
{kind=link}
{kind=link}