Expression and assembly of HIV-1 Gag cleavage-deficient mutants
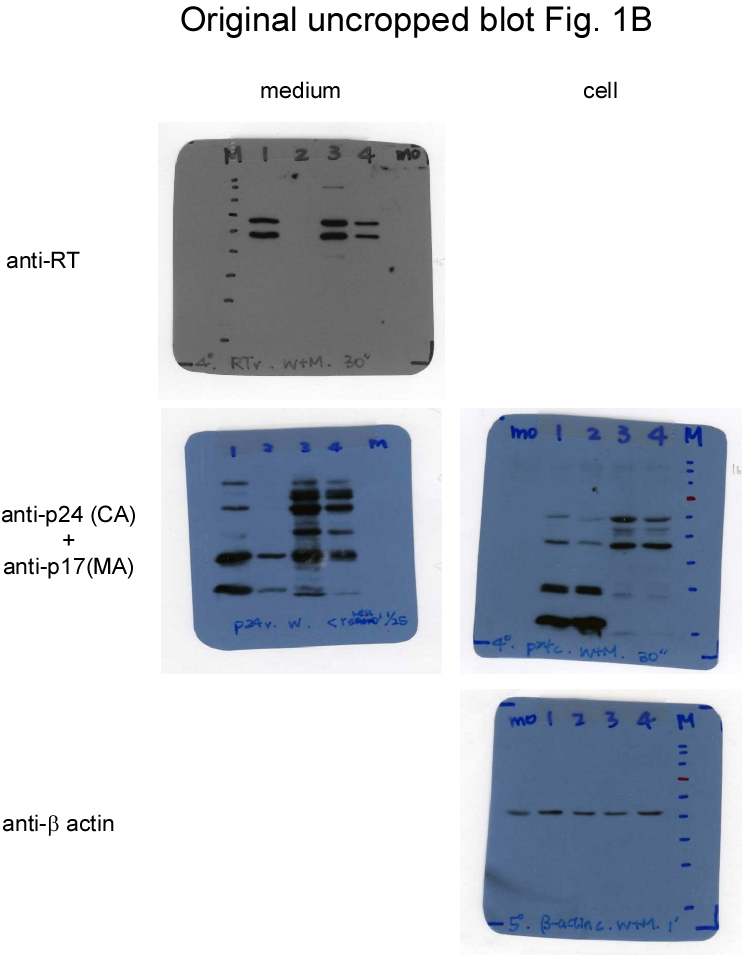
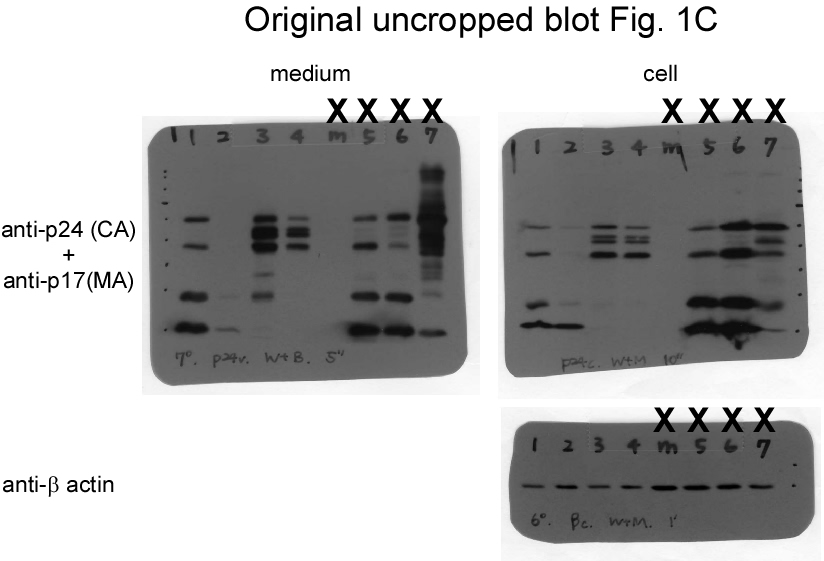
To test the effects of Gag cleavage on virus assembly and Gag-Pol packaging, a mutant (designated CSM) with amino acid substitution mutations was engineered to block PR cleaving at p2/NC and MA/CA (Fig. 1a). Since Gag-Pol incorporation depends on interaction with Pr55gag, and since Gag cleavage efficiency can affect virus particle production, we used medium-associated Gag—specifically, Gag precursor Pr55gag, incompletely processed p41gag, and/or mature p24gag—as a virus production marker. Data for transient expression in 293T cells indicate significant reductions in the CSM cellular products p24gag and p17gag compared to those of a wt (Fig. 1b, lower panel, lane 4 vs. lane 2; supplementary Fig. 1b), confirming PR cleavage inhibition at the p2/NC and MA/CA junctions. The observation that CSM exhibited virus-associated p24gag at a level comparable to that of the wt is likely due, at least in part, to western blot overexposure (Fig. 1b, middle panel, lane 4 vs. lane 2). In a repeat experiment we found that the level of CSM virus-associated p24gag was noticeably lower than that of the wt (Fig. 1c, upper panel, lane 3 vs. lane 1; supplementary Fig. 1c). As a result of this blocking, substantial amounts of intermediate or incompletely processed Gag were detected in CSM supernatant and cellular samples. A band migrating slightly faster than wt p41gag (MA-CA-p2) likely corresponded to the MA-CA molecular weight (Figs. 1b and 1c, asterisks). Bands migrating to positions between Pr55gag and p41gag likely corresponded to the MA-CA-p2-NC(-p1) molecular weight (Fig. 1b, upper panel, lanes 4-5, and Fig. 1c, lanes 3-4). Some of the aberrant p24gag-associated products may have resulted from cleavages at cryptic sites when MA/CA- and p2/NC-accessible sites were blocked. These partial Gag cleavage products might assemble and release as VLPs, which would be consistent with prior findings that Gag lacking p6 and/or NC domains is still capable of assembly and release as VLPs in 293T cells [34, 35].
Effects of enhanced Gag cleavage on virus assembly and Pol packaging
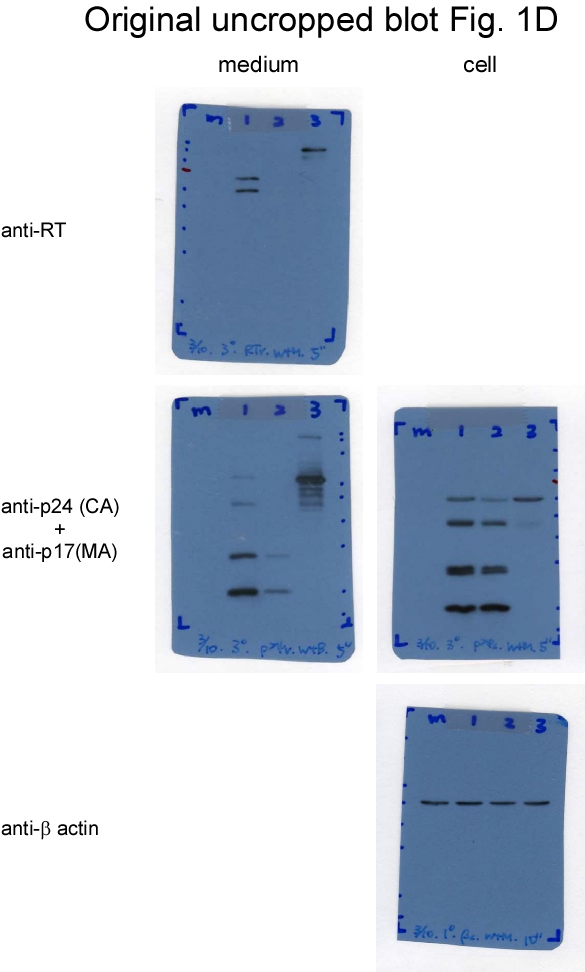
We posited that Gag-Pol dimerization facilitated by EFV might enhance Gag-Pol autocleaving, with the possibility of reduced virus-associated RT or Gag-Pol levels resulting (at least in part) from premature Gag cleavage due to the enhanced autocleaving effect. Treatment with 1 µM or 5 µM EFV was found to significantly reduce virus yields as a result of enhanced Pr55gag cleavage [16, 29]. Accordingly, CSM virus particles might contain higher levels of RT or Gag-Pol when PR activation is enhanced. We treated wt and CSM transfectants with 1 µm EFV to test this possibility. Consistent with previously reported results [30], EFV treatment triggered a significant decrease in wt virus yield due to enhanced Pr55gag cleavage efficiency (Figs. 1b. lane 3, 1c, lane 2 and 1d, lane 3, supplementary Figs. 1b-d). The Pr55gag cleavage enhancement effect from EFV is PR activity-dependent, as evidenced by the detection of substantial amounts of Pr55gag particles following treatment with an HIV-1 PR inhibitor (Fig. 1d, lane 4). In contrast, CSM virus yields were only moderately reduced following EFV treatment (Fig. 1b, lane 4 vs lane 5), suggesting that blocking PR to induce cleavages at MA/CA and p2/NC can counteract the negative EFV effect on virus production. Similar to the decrease in virus production, virus-associated RT levels also declined following EFV treatment.
Blocking Pr55 gag cleavage mitigates negative impacts of Gag-Pol overexpression on virus yields and RT packaging
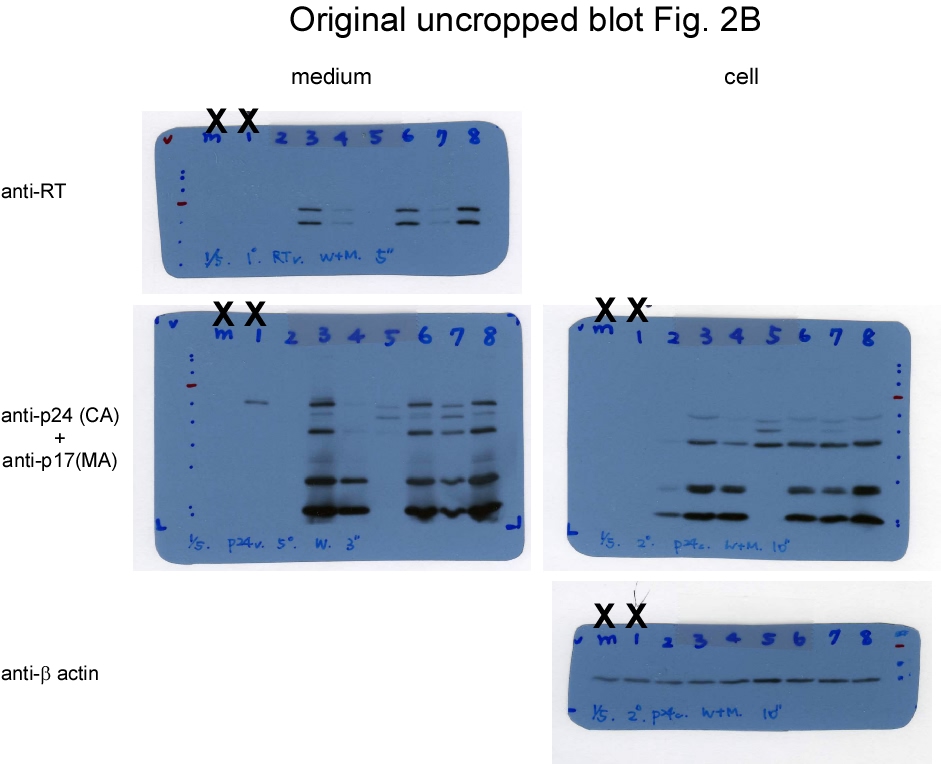
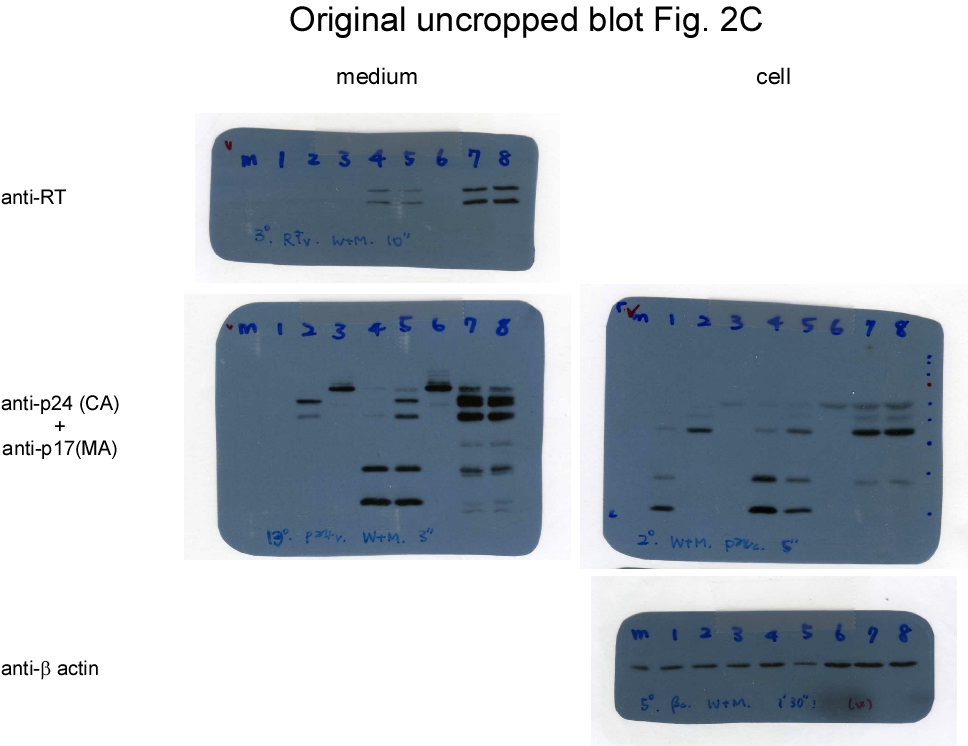
Since PR can access cleavage sites on Pr55gag and Gag-Pol, and since reduced virus yields due to enhanced Pr55gag cleavage are frequently associated with reduced Gag-Pol packaging, it is difficult to assess the impacts of Gag cleavage inhibition on Gag-Pol packaging when both Pr55gag and Gag-Pol are expressed from the same plasmid, as observed in the CSM case. To address this challenge, we inserted the p2/NC and MA/CA cleavage site mutations into a Gag-Pol expression vector GPfs, and co-transfected the resulting construct (designated CSMfs) with either a wt- or CSM-containing Gag expression vector (Fig. 2a). The GPfs vector contains pol and gag in the same reading frame, resulting in the expression of Pr160gag−pol only [15]. Our data indicate that virus-associated RT, p24gag, and p17gag were readily detected when CSMfs or GPfs were co-transfected with a Pr55gag-expression vector (Gag) at a DNA ratio of 1:10 (Fig. 2b, lanes 2 and 5; supplementary Fig. 2b). This suggests that Gag cleavage blocking within Gag-Pol did not exert significant impacts on the ability of Gag-Pol to be incorporated into Pr55gag particles in trans when the Gag-Pol/Pr55gag expression ratio was close to matching physiological conditions. Unsurprisingly, noticeable reductions in virus-associated Gag and RT resulted when a Pr55gag-expressing vector was co-transfected with equal amounts of GPfs or CSMfs plasmid DNA (Fig. 2b lanes 3 and 6 and Fig. 2c lanes 5-6).
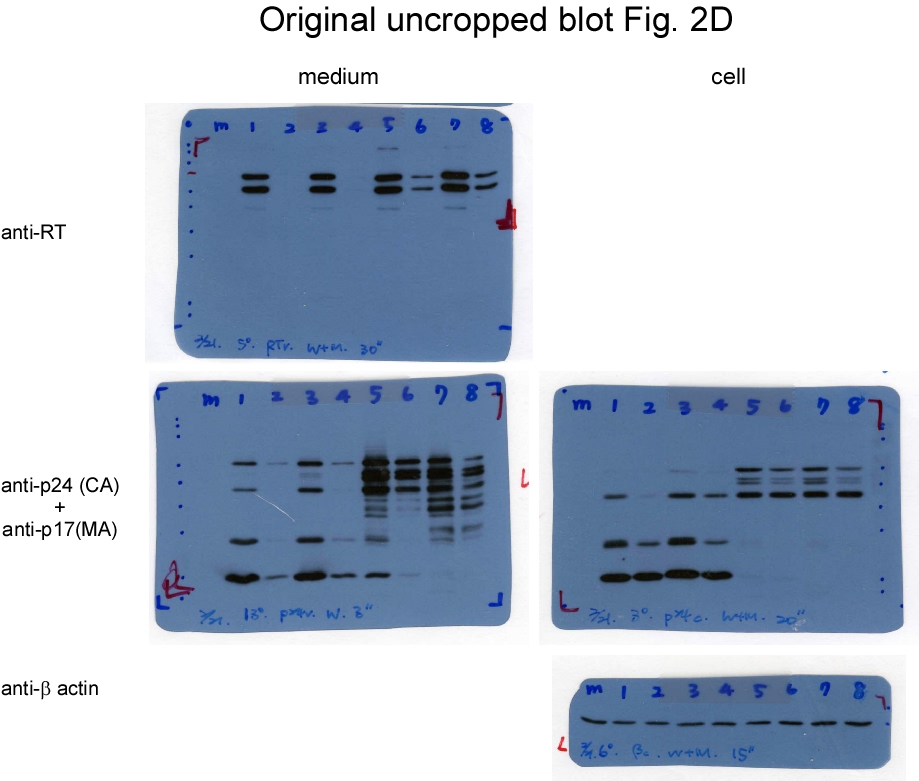
These data are in agreement with previously reported findings that Gag-Pol overexpression markedly reduces VLP yields. We noted that either GPfs or CSMfs overexpression with wt Gag (Pr55gag) resulted in barely detectable virus-associated RT accompanied by a reduction in virus-associated Gag. In contrast, virus-associated RT became readily detected with increasing virus-associated Gag when GPfs or CSMfs was co-expressed with CSMGag under the same condition (Fig. 2c, lanes 8-9 vs. lanes 5-6; supplementary Fig. 2c). No Gag products were detected when GPfs was expressed alone. However, p24gag-associated Gag products (likely corresponding to MA-CA-p2-NC and MA-CA) were detected in CSMfs transfectant supernatants (Fig. 2c middle panel, lane 2 vs. lane 3). This suggests that blocking Gag-Pol N-terminal Gag cleavage may yield incompletely processed Gag that is capable of forming VLPs, as observed in the CSM case (Fig. 1). Similar to the Gag-Pol overexpression scenario (Fig. 2c), virus yields decreased when PR activation was enhanced via EFV treatment (Fig. 2d; supplementary Fig. 2d). However, we did not observe significant differences between GPfs and CSMfs in terms of more efficient incorporation into wt Gag or CSMGag particles.
Effects Of Pr Activity Inhibition On Pol Packaging
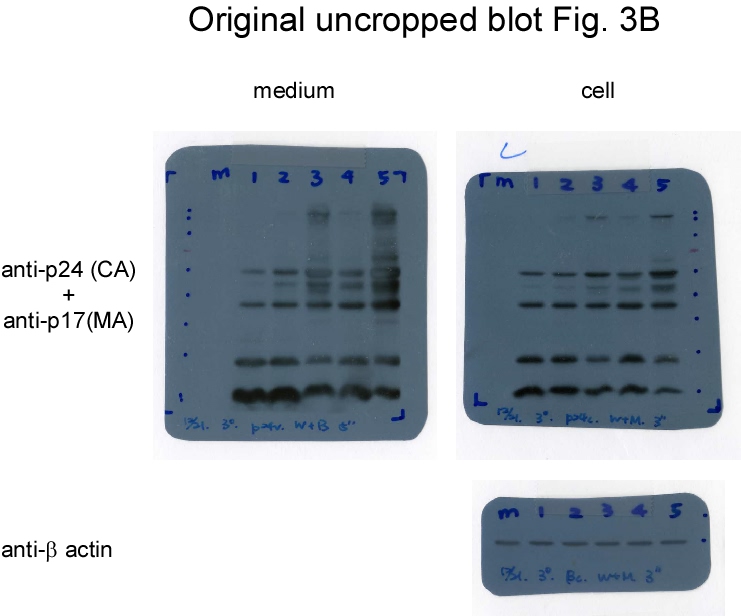
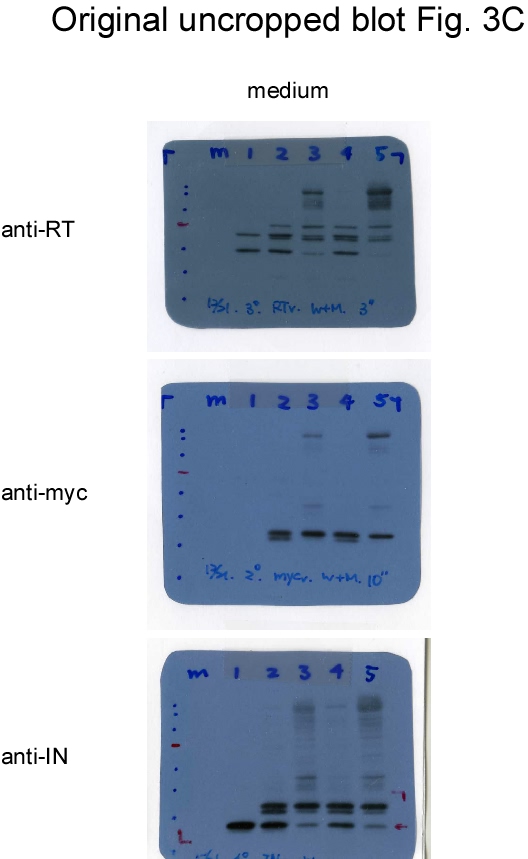
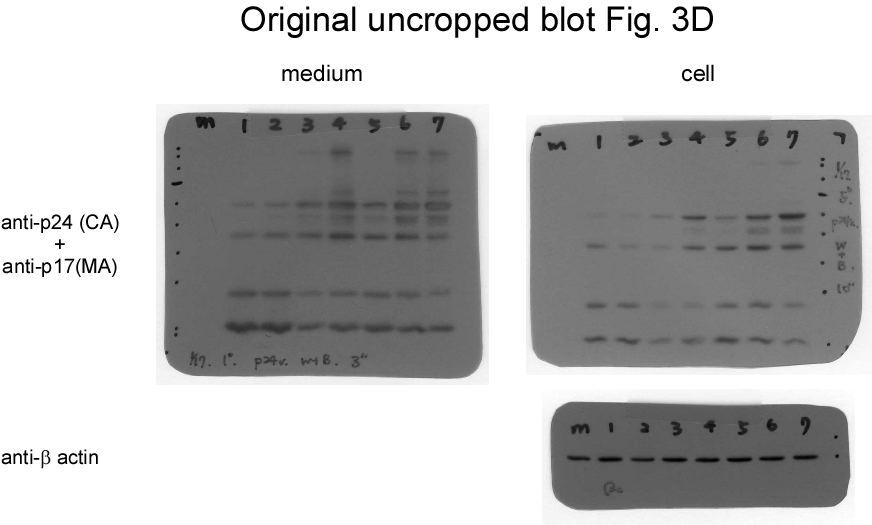
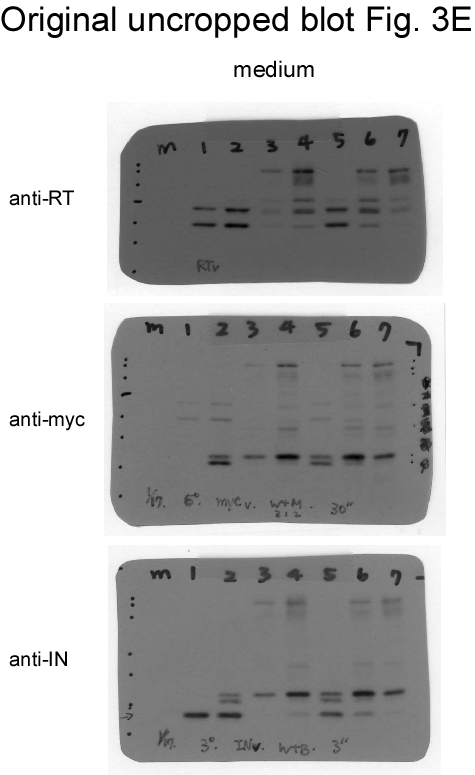
If Gag-Pol autocleaving occurs during or prior to viral incorporation, there is a possibility that PR-defective Gag-Pol may be packaged more efficiently than its PR-active counterpart. To test this possibility, PR-defective Gag-Pol with or without the CSM mutation was co-expressed with a wt. To distinguish PR-defective Gag-Pol from wt Gag-Pol, a Myc epitope was tagged at the C-terminus of PR-defective Gag-Pol and designated fsDmyc or CSMfsDmyc (Fig. 3a). Transient co-expression results indicate that increases in co-transfected amounts of these PR-defective Gag-Pol expression vectors led to increases in Gag and Gag-Pol precursors associated with moderate decreases of RT66/51 and integrase (IN) in supernatant samples (Figs. 3b-c, lanes 3-6, and Figs. 3d-e, lanes 3-8; supplementary Figs. 3b-e). These findings suggest that due to overexpression, PR-defective Gag-Pol is packaged at a higher frequency than wt Gag-Pol, and that the defective PR domain interferes with wt PR activity, resulting in the inefficient processing of Pr55gag and Gag-Pol. Virus-associated IN and IN-myc levels were roughly equal when co-transfected PR-defective Gag-Pol plasmids were reduced to near-wt Gag-Pol expression levels (Fig. 3c, lanes 3 and 5). Combined, these data suggest that wt Gag-Pol is capable of mediating the processing of incorporated PR-defective Gag-Pol in trans, and that PR-defective Gag-Pol (with or without CSM) does not exhibit greater packaging efficiency compared to wt Gag-Pol when the two are co-expressed at approximately equal levels.
Effects Of Enhanced Gag-pol Autocleavage On Pol Packaging
In a previous report we described a Dp6*PR construct containing a duplicate p6*-PR with an inactivated proximal PR copy exhibiting a wt virus processing profile [31]. However, in that study virus-associated Gag was barely detectable when a leucine zipper (LZ) dimerization motif was substituted for the p6* and placed adjacent to the active PR. The severe assembly defect of this mutant (designated DWzPR) resulted from enhanced PR activation due to an LZ substitution for the deleted p6*, but a DWz/PR construct containing four C-terminal p6* residues in the deleted p6* region was found to counteract the LZ enhancement of PR activation [19]. We hypothesized that enhanced Gag-Pol autocleavage triggered by LZ may lead to reduced Pol viral incorporation, and that preventing Gag cleavage from Gag-Pol might retain that incorporation at the same level.
To test this possibility, a CSM mutation was cloned into DWzPR and DWz/PR, respectively yielding CSM/DWzPR and CSM/DWz/PR (Fig. 4a). Transient expression results indicate that DWzPR produced barely detectable virus-associated Gag, while DWz/PR virus-associated Gag products were readily detectable (Fig. 4b; supplementary Fig. 4b); this is consistent with previous reports [19, 32]. We found that compared to the wt transfectant, cellular Pr55gag was barely detectable in DWzPR and DWz/PR transfectants (Fig. 4b lower panel, lanes 2-4), indicating enhanced PR-mediated Gag cleavage efficiency. Both DWzPR and DWz/PR exhibited barely detectable or noticeably reduced virus-associated RT compared to their CSM-containing counterparts (Fig. 4b upper panel, lanes 3-4 vs. lanes 6-7, and Fig. 4c).
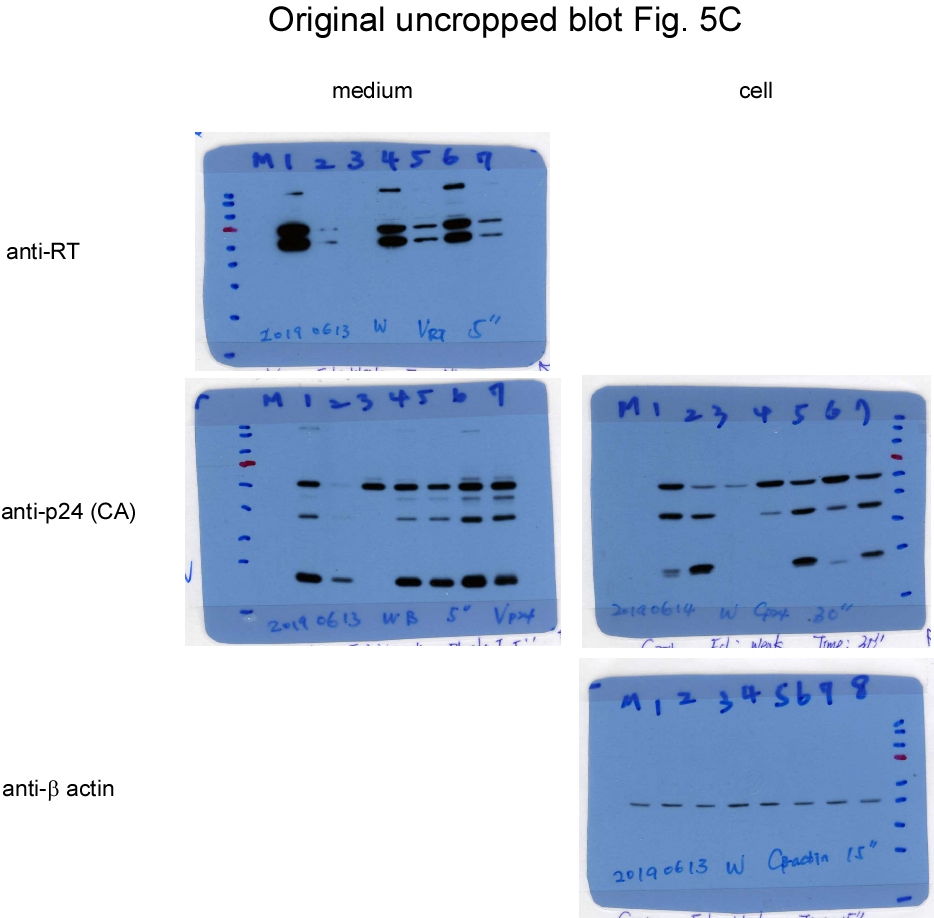
The results shown in Figure 4 suggest that blocking Gag cleavage can partly counteract the reduced Pol packaging incurred by the LZ replacement of p6*. However, Pol or Gag-Pol packaging is primarily determined by their respective ability to associate with Pr55gag. In addition to reducing VLPs, enhanced PR activation may reduce Gag-Pol viral packaging as a result of enhanced Gag-Pol autocleavage. Since it is difficult to assess the impact of enhanced Gag-Pol autocleavage incurred by LZ on Gag-Pol packaging when Gag cleavage sites within Pr55gag and Gag-Pol are blocked, we assessed the degree of DWZPR and DWz/PR Gag-Pol incorporation into wt Pr55gag particles. After cloning DWzPR and DWz/PR with or without the CSM mutation into the GPfs backbone (Fig. 5a), each resultant construct was co-transfected with the Pr55gag expression vector. Our results indicate that the leucine zipper replacement of p6* significantly reduced RT-associated products in virus particles (Fig. 5b upper panel, lanes 3-4 and 6-7 and Fig. 5d; supplementary Fig. 5b). However, virus-associated RT levels in DWzPRfs and DWz/PRfs did not increase significantly when Gag cleavage sites were blocked (Fig. 5b lane 3 vs. lane 6 and lane 4 vs. lane 7; Fig. 5d). As an additional test, Pr55gag was co-expressed with a W420A-containing GPfs construct with or without the CSM mutation. W402A (with an alanine substitution of the RT codon W402) was found to be severely assembly-defective due to enhanced PR activation triggered by the W402A mutation [33]. Consistent with Chiang et al.’s results, W402A exhibited markedly reduced VLP yields compared to the wt (Fig. 5c, lanes 2-3; supplementary Fig. 5c) and reduced Pol incorporation into Pr55gag particles, regardless of whether or not the Gag-Pol contained CSM (Fig. 5c upper panel, lanes 5-8).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}