Here, we investigated the comparative anatomy of fleshy diaspores in South American Ephedra, an aspect of great relevance to their reproductive biology. We found that (1) Fleshy and papery seed cone bracts differ anatomically since early developmental stages, the former having mucilage chlorenchyma, (2) Mucilage underlies the gelatinous texture of fleshy seed cone bracts; (3) Cellulosic fibers are found in bracts, where they likely function as apoplastic water “pipes”, and (4) Young leaves and bracts share the same type of mucilage tissue in all fleshy species studied. We therefore propose that bract fleshiness is a novel differentiation program repurposed from mucilage tissue present in leaves, which develops similarly in all species. Additionally, we propose that the diversity of bract morphology and anatomy found in Ephedra likely evolved via heterochronic processes involving intra-individual changes in the timing of leaf developmental events.
Fleshy and papery seed cone bracts differ anatomically since pollination stage
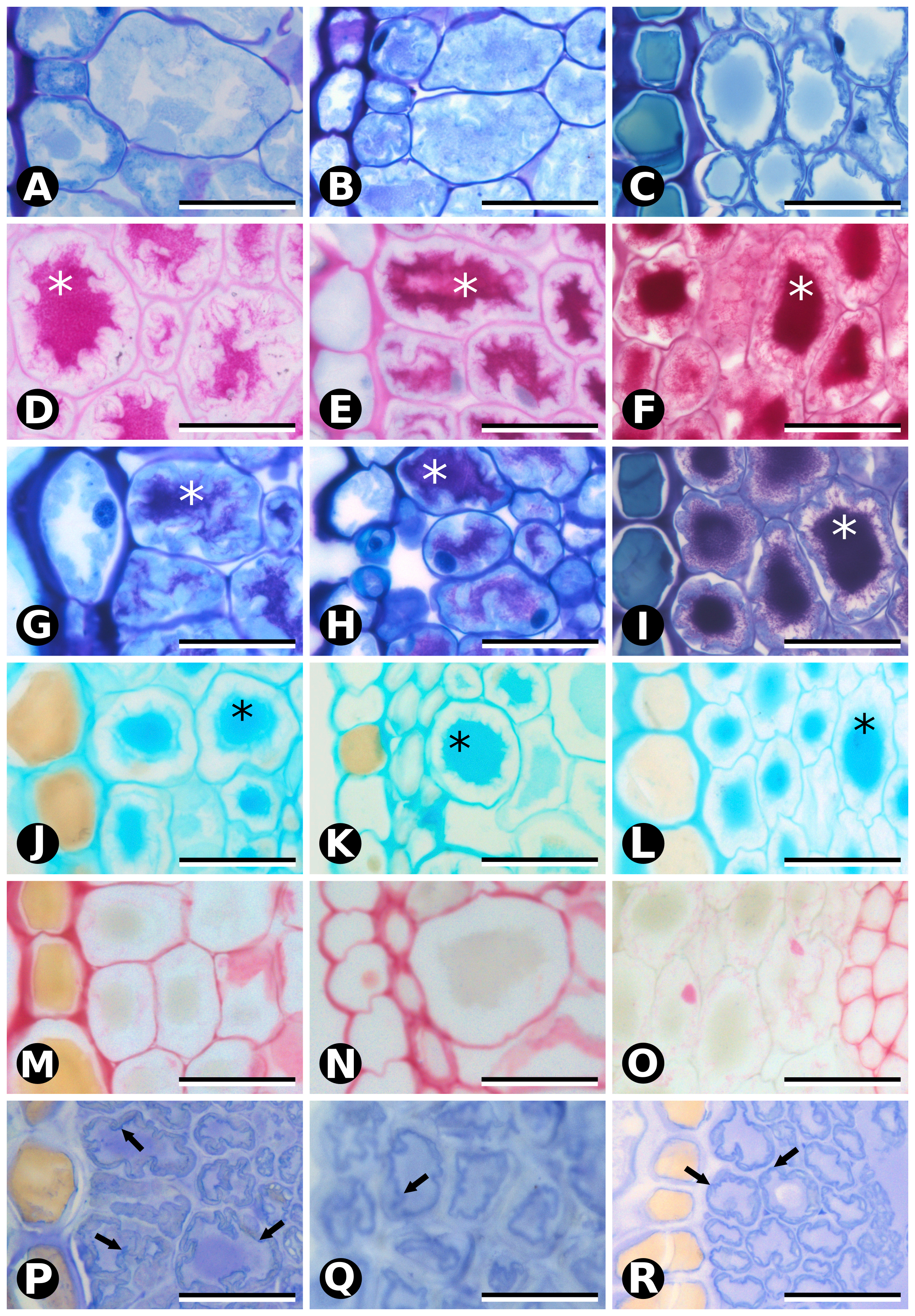
Our ontogenetic observations and biochemical assays provide evidence that seed cone bracts from fleshy species develop previously undescribed, highly specialized mucilage chlorenchyma already discernible at the early stages of pollination, and not found in dry bracts at the same stage (Fig. 8A). This mucilage chlorenchyma can also be found in the bracts of pollen cones of the fleshy species, except for E. chilensis, where tanniniferous cells are found instead (Fig. 8B).
Mucilage accounts for the gelatinous texture of Ephedra fleshy seed cone bracts
Seed cone bracts of E. chilensis, E. triandra and E. tweediana have uniform, mucilage chlorenchyma mesophyll at the pollination stage (stage 1) that becomes distinctly red and fleshy at the seed dispersal stage (stage 3). Our histochemical assays identified mucilage as the substance responsible for the gelatinous texture of the mature red bracts of E. chilensis, E. triandra and E. tweediana seed cones. Mucilage comprises a family of highly branched, polymerized carbohydrates including L-arabinose, D-galactose, L-rhamnose, D-xylose, and galacturonic acid in various proportions that may also contain glycoproteins and other compounds such as tannins [14]. Whereas we found mucilage inside the cytoplasm of cells of a specialized type of parenchyma in Ephedra, mucilage is only found outside the cell, or in specialized cells known as idioblasts in angiosperms [15–17]. While our analysis was not able to detect the type and location of the pigment/s responsible for the red color at later developmental stages (stages 2 and 3), anthocyanins have been reported in fleshy red gymnosperm bracts and seeds [18], and rhodoxanthin is another good candidate found in Ephedra monosperma leaves [19].
Cellulosic fibers as apoplastic “water pipes”, an adaptation to arid environments
Another novel observation was the presence of cellulosic fibers in Ephedra cone bracts and leaves. Cellulosic fibers are similar to G-fibers in overall morphology, both are an infrequent cell type in gymnosperms only known otherwise from the wood of certain conifers [20], roots of cycads, and vegetative organs of Gnetales [21–25]. However, they have differences in structure and function: G-fibers or gelatinous fibers (also known as tension fibers) are long, fusiform, unbranched cells with lignified primary and secondary walls and a non-lignified innermost wall or gelatinous layer (Sg, or G-layer), composed mostly of lamellate cellulose [24]. Cellulosic fibers are also long and fusiform, but they are usually branched, with non-lignified primary and secondary walls and a tertiary wall of microcrystalline cellulose. In this study, we adopt “cellulosic fibers” to distinguish a particular type of fibers without lignin from the typical g-fibers, which usually differentiate in response to external forces, such as gravity or wind [24]. The specialized cell wall structure of G-fibers allows them to contract and produce counteracting tensile forces to reorient organs [26], as in Gnetales stems and roots [23]. Cellulosic fibers are only known for Gnetales within gymnosperms: as hypodermal fibers in Welwitschia leaves [21, 27], as cellulose fibers in Gnetum leaves [22], and also misidentified as g-fibers in the cortex of Ephedra stems [25]. In Gnetum leaves, cellulosic fibers perform a supplementary apoplastic water transport function to compensate for the low density of leaf veins [28], rather than the typical mechanical function. A water transport function is also compatible with a putative role in protecting axillary buds (leaves), and reproductive strobili (pollen and seed cone bracts) against desiccation early in development, especially relevant in the semi-arid to arid environments inhabited by Ephedra in general [13], and the South American species in particular (Fig. 1). We therefore propose that this putative function of cellulosic fibers in leaves has been repurposed as a supplementary water supply system for the development of fleshy seed cone bracts that facilitate seed dispersal.
Shared leaf anatomy across species differs in the timing of maturation
Mature Ephedra leaves are typically reduced, scale-like, dry and membranous, and whither early; with photosynthetic function being transferred to the green stems [29]. These leaves consist of an epidermis collapsed to different degrees and lacking mesophyll, except in E. altissima, which develops linear leaves with little anatomical differentiation [29]. Our histological observations on leaves confirmed their reduced structure, while highlighting that they still contain distinct types of mesophyll cells at maturity: adaxial cellulosic fibers, abaxial tanniniferous cells, transfusion tissue and vascular tissue. This mature leaf anatomy is similar between male and female individuals of the same species, and between species with fleshy or papery seed cone bracts (Fig. 8C-D). Young (second node) leaves of fleshy species develop adaxial compact parenchyma and abaxial mucilage chlorenchyma prior to differentiating cellulosic fibers or tanniniferous cells, respectively. The transformation of chlorenchyma cells into tanniniferous cells is consistent with tannins being synthesized in chloroplast-derived organelles (tanossomes) [30, 31], and accumulating in the vacuole. Leaves of fleshy species appear to differ, however, in the timing of maturation (Fig. 9). Young leaves (second node) of the papery species E. multiflora already have mature anatomy, although we could not analyze leaves in the bud to verify whether mucilage chlorenchyma precedes tanniniferous cells (as in leaves of fleshy species), or whether they never form. Mucilage chlorenchyma in E. multiflora leaf primordia would account for a much earlier differentiation of leaf anatomy, while its absence would indicate a profound difference in foliar structure between fleshy and papery species. Despite being non-photosynthetic and highly reduced, mature Ephedra leaves are not completely dead structures, since tanniniferous cells are alive and cellulosic fibers can potentially continue to supply water via apoplastic transport. Considering all the evidence, mature leaves more likely perform a protective function towards axillary buds, against dehydration, heat, UV radiation [32] and herbivory [33].
Bract fleshiness is a novel differentiation program repurposed from leaves
Mucilage cells found in fleshy seed cone bracts are also found in young leaves, but while they turn into tanniniferous cells in leaves, they instead produce red pigment and expand into fleshy structures in seed cone bracts, presumably by increasing water intake. We propose that the terminal “leaf differentiation program” (mucilage cells becoming tanniniferous cells) fulfills a protective function that has been repurposed into a “fleshy bract differentiation program” (mucilage cells dividing, expanding, becoming fleshy and red) for a novel, seed dispersal role. Papery bracts of E. multiflora pollen and seed cones follow the differentiation program of leaves, as do pollen cone bracts of E. chilensis (Fig. 8).
Changes in the timing of leaf development drive seed cone bract diversification
Our results also revealed that reproductive bracts are anatomically more diverse than vegetative leaves in Ephedra, not due to new cell types, but to novel combinations enabling alternative differentiation pathways that depart from baseline leaf development. When comparing and contrasting bract and leaf anatomy (Fig. 8A-B vs. 8C-D), the structure of pollen cone bracts can be a) the same as seed cone bracts and leaves (Ephedra multiflora), b) different from seed cone bracts but similar to leaves (E. chilensis), or c) different from seed cone bracts and leaves (E. tweediana and E. triandra). Sexual dimorphism in cone bracts in the “fleshy species'' is presumably related to their respective functions in pollen protection vs. animal seed dispersal via independent selective forces. At first sight, this reasoning appears to fail to explain why pollen and seed cone bracts of E. multiflora would be similar, until we consider the plant in its natural habitat: the selective pressures for dispersal by wind at high elevation desolate environments, where animals are scarce and wind speeds are high (Fig. 1G and Additional File 2).
From an ontogenetic perspective, bract diversity can be explained based on changes in the timing of leaf development. On the one hand, pollen and seed cone bract morphology resembles juvenile leaf shape with reduced hyaline margins in “fleshy species” (E. chilensis, E. triandra and E. tweediana) (Fig. 9A-D). The increase in cell layers between stages 2 and 3 of fleshy seed cone bracts leading to their maturity can be interpreted as a juvenile leaf developmental stage. Mesophyll of E. chilensis pollen cone bracts resembles mature leaf mesophyll (Fig. 9E and I), while that of E. triandra and E. tweediana matches young leaf anatomy (Fig. 9D and H). On the other hand, papery bracts of E. multiflora reflect senescent, or hypermature leaf features, both in their mesophyll anatomy and in the wide hyaline margins (Fig. 9F, J and K). Thus, bract structure in “fleshy species” may be interpreted as based upon juvenile stages of leaf development, while that of “papery species” would result from the extension or acceleration of the mature leaf development program.
Heterochrony represents changes in the timing of developmental events leading to phenotypic variation between the ancestor and its descendants [34, 35]. While different types of heterochrony have been described: interspecific vs. intraspecific [36], growth vs. sequence [37], transcriptional vs. metabolic vs. cellular [38], all of them are framed in an evolutionary context (between ancestor and descendants), or among individuals of the same species. In our study, the proposed changes in the timing of developmental events would be modifying homologous structures on the same individual, since leaves and bracts are considered serial (iterative) homologous structures [39], yet the adoption of heterochrony for serially homologous structures has not been fully developed [40, 41]. In the meantime, ancestral character reconstruction at the genus level would further test the potential role of heterochrony in cone bract evolution in Ephedra.
Evolutionary considerations
Overall, bract anatomy was more nuanced than expected: remarkably similar between sister species Ephedra tweediana and E. triandra (for both pollen and seed cone bracts), yet markedly different from E. chilensis pollen cone bracts (sister species of E. tweediana + E. triandra) [13]. “Fleshy” seed cone bracts are considered the ancestral state for the South American clade, while dry papery bracts have presumably evolved independently multiple times in Ephedra [13], yet only once in the South American clade, in the branch leading to E. multiflora and E. boelckei [12, 13]. Here, we contribute anatomical and histological evidence for Ephedra cone bracts that suggests that the ancestral fleshy character state may represent a novel type of bract differentiation built upon young leaf anatomy, while the derived dry, papery bract state is based on an existing differentiation pattern found in mature vegetative leaves. Additionally, we propose that cellulosic fibers observed here in Ephedra leaves and bracts, and also found in Gnetum [22] and Welwitschia [21, 27], may constitute a previously overlooked valuable synapomorphy for the order Gnetales.
{kind=link}