Phenotypic and Physiological analysis of two varieties
WL is known to induce chlorosis and early senescence of leaves [26]; Firstly, we detected the chlorophyll concentration in expanded flag leaves of WL-tolerant variety XM55 and WL-sensitive variety YM158 by measuring SPAD (soil-plant analysis development) at the anthesis stage. The SPAD value of XM55 was higher than that of YM158 during 0-7 days, and it was less than or equal to that of YM158 during 7-21 d under WL (Fig. 1A). However, the SPAD value of XM55 was higher than that of YM158 between 0-21 days under normal conditions (CK) (Fig. 1A). The SPAD value of XM55 under normal conditions decreased below that of WL treated XM55 after 7 d, whereas it was decreased below that of YM158 under CK at 5 d (Fig. 1A). The reductions of SPAD in XM55 from 0 to 7 d, 7 to 14 d, and 14 to 21 d under WL were 2.7%, 4.2%, and 7.8%, whereas they were 4.7%, 6.9%, and 13.4% in YM158, respectively.
Soil WL causes serious hypoxia in plant roots, obstructs root growth and development, decreases root activity, and decreases root water permeability; this affects plant water uptake and transpiration rate, thereby leading to water deficit in plants and alterations in the above-ground distribution of water [26, 27]. We also measured the above-ground water contents in the two varieties. Under WL, the water contents in flag leaves, ears, and stem and sheath were significantly higher in XM55 than in YM158 from 7 to 21 d, whereas this pattern occurred from 14 to 21 d under CK (Fig. 1B, 1C, 1D)
WL at elongation or post-anthesis is known to affect grain yield, as well as accumulation and remobilization of dry matter in wheat [27]. We measured the changes of aboveground dry matter accumulation (DMA), yield, and yield-related traits of the two varieties. WL had different effects on XM55 and YM158. The DMA at anthesis (DMA1) before WL were roughly similar between XM55 and YM158, but under WL, DMA values were decreased by 12.5% and 20.5% in XM55 and YM158 relative to the CK control at the mature stage, respectively (Table 1). At the same time, kernels per spike, 1000-kernel weight, grain yield weight, and harvest index were decreased under WL by 3.3, 18.1, 26.2 and 15.9% in XM55, and by 10.8, 36.2, 36.8 and 21.8% in YM158 relative to their CK control values, respectively (Table 1). Clearly, WL had greater effects on YM158 instead of XM55, especially the 1000-kernel weight and grain yield weight. Overall, it could be inferred that XM55 showed better WL tolerant than YM158.
Waterlogging Induced Proteome Change in XM55 and YM158
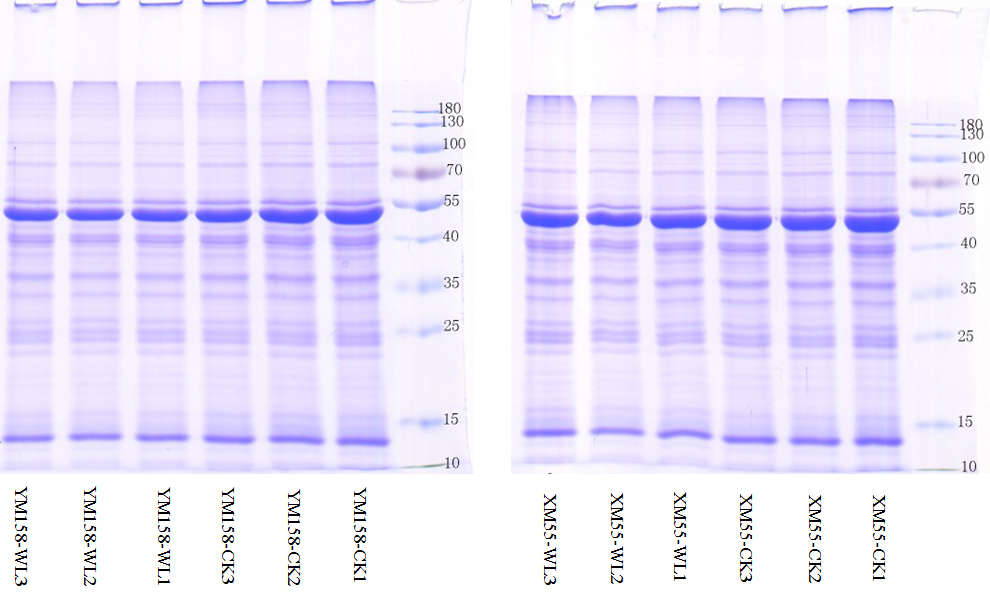
To further explore the molecular mechanisms that mediate different responses to WL, iTRAQ method was used to analyze proteome changes in flag leaf of both cultivars. After protein extraction, enzyme digestion, iTRAQ labeling, equal mixing and SCX pre-separation, all samples were subjected to LC-MS/MS in three independent replicates. In the present study, a total of 1,087,846 spectra were detected, among which, 37,952 could be matched and 55,206 were unique spectra, and 37,985 peptides could be identified with 19,279 being unique peptides, and 7710 proteins were identified (Fig. 2A); the proteins identified in the flag leaf of the XM55 and YM158 plants were supported by unique peptides. Of those proteins, 54.0% (4,164) were inferred from more than three unique peptides (Fig. 2B).
Pairwise comparison of protein profiling in XM55 and YM158 under WL
To identify differentially expressed proteins (DEPs) between the two cultivars in response to WL, proteins with more than a 1.2-fold change in abundance (p < 0.05) between XM55 and YM158 under WL and CK were investigated. Based on this criterion, 23 DEPs (14 up-regulated and 9 down-regulated) showed differential expressed between XM55 and YM158 under WL (XM55-WL/YM158-WL), and 52 DEPs (31 up-regulated and 21 down-regulated) were differently expressed between XM55 and YM158 under CK (XM55-CK/YM158-CK) (Fig. 3). At the same time, 7 proteins (i.e., TRIAE_CS42_2BL_TGACv1_130584_AA0414140.1, TRIAE_CS42_2BL_TGACv1_131439_AA0427700.2, TRIAE_CS42_4BL_TGACv1_321826_AA1065960.1, TRIAE_CS42_2BL_TGACv1_132610_AA0438610.1, TRIAE_CS42_6BL_TGACv1_503168_AA1627380.1, TRIAE_CS42_6BL_TGACv1_503168_AA1627380.2, TRIAE_CS42_6BL_TGACv1_503168_AA1627380.3) were differentially accumulated in both (XM55-WL/YM158-WL) and (XM55-CK/YM158-CK), which might indicate cultivar specific protein accumulation irrespective of WL treatment (Fig. 3, Table 2, Table S1). Excluding these 7 overlapping DEPs, a total of 16 DEPs were remained between XM55 and YM158 under WL (Fig. 3B); of these, 11 DEPs were up-regulated, including members of Fe-S cluster assembly factor, heat shock cognate 70 kDa protein, GTP-binding protein SAR1A-like, and CBS domain-containing protein, respectively. The 5 down-regulated proteins were photosystem II reaction center protein H, carotenoid 9,10 (9',10')-cleavage dioxygenase-like, psbP-like protein 1, and mitochondrial ATPase inhibitor (Table 2).
Proteomic dynamics in XM55 and YM158 between WL and CK
The DEPs in XM55 and YM158 between WL and CK were identified. There were 84 DEPs (35 up-regulated and 49 down-regulated) between XM55-WL and XM55-CK, and 59 DEPs (13 up-regulated and 46 down-regulated) between YM158-WL and YM158-CK (Fig. 3, Table S2, Table S3). Most proteins responsive to WL were specific to XM55 or YM158. However, 9 proteins were differentially expressed in both XM55-WL/XM-CK and YM158-WL/YM158-CK (Fig. 3, Table S2, Table S3), which might be WL responsible proteins with non-cultivar specificity. These proteins were 3-isopropylmalate dehydratase large subunit, solanesyl-diphosphate synthase 2, DEAD-box ATP-dependent RNA helicase 3, and three predicted or Uncharacterized proteins. 3-isopropylmalate dehydratase catalyses, the stereo-specific isomerization of 2-isopropylmalate and 3-isopropylmalate participate in the biosynthesis of leucine.
Functional Categorization, GO and KEGG pathway Enrichment Analysis of the DEPs
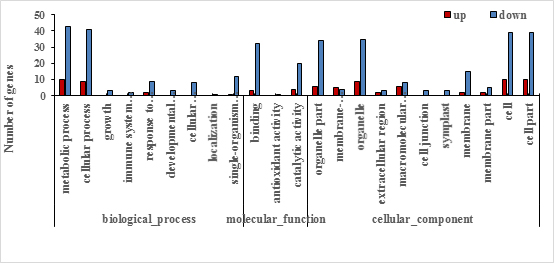
The functional information of all differentially accumulated proteins in Fig. 3 were obtained by searching against the UniProt-GOA database, which were assigned to three categories based on GO annotation, that is, cellular compartment, biological process, and molecular function. The differentially expressed proteins among XM55 and YM158 under WL belonged to eight biological processes, 11 cellular compartments, and two different molecular functions (Fig. 4, Table S4). In terms of biological processes, metabolic process, cellular process and cellular component organization or biogenesis were the three major groups. It was suggested that the DEPs may be involved in primary metabolic processes, and these impart differential WL tolerances to XM55 and YM158. Cell, cell part, and membrane-enclosed lumen were the top three cellular compartments, implying that various changes in cell structure had effects on tolerance to WL among different varieties. Binding was the major molecular functional groups, and a small amount of differentially accumulated proteins were involved in catalytic activity, which showed that protein binding affects tolerance to WL.
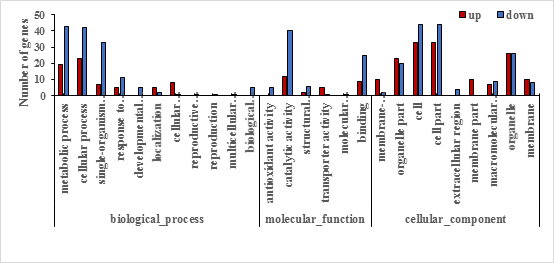
The differentially expressed proteins among XM55 or YM158 under WL and CK belonged to 11 or 8 biological processes, 9 or 11 cellular compartments, and 6 or 3 molecular functions (Fig. S1, Fig. S2, Table S4), respectively. Metabolic process, cellular process, and single-organism process were both the three major biological processes. Cell, cell part, and organelle were both the top three cellular compartments. Catalytic activity and binding were both the two-major molecular functional groups. Those results indicated that primary metabolic processes, cell structure, and catalytic activity were generally affected by WL regardless of cultivar tolerance.
To characterize the functional consequences of the differentially expressed proteins associated with WL, the enriched pathways were assigned based on KEGG terms. The results indicated that the proteins related to terpenoid backbone biosynthesis, amino sugar and nucleotide sugar metabolism, and fructose and mannose metabolism were affected by WL in XM55, whereas terpenoid backbone biosynthesis and fatty acid biosynthesis were affected in YM158. Tuberculosis and RNA degradation were affected by WL both in XM55 and YM158 (Table S5).
Correlation of differentially accumulated protein with mRNA Expression
To verify the correlation between the expression levels of the differentially expressed proteins and their mRNAs, the mRNA expression levels of 28 differentially expressed proteins were analyzed using qRT-PCR method (Table S6). Among them, 16 genes exhibited consistent expression patterns with their proteins, whereas 12 showed discrepancies between protein accumulation and mRNA expression (Fig. 5). The discrepancy between protein accumulation and mRNA expressions might be ascribed to translational and posttranslational regulatory processes or feedback loops between the processes of mRNA translation and protein degradation [28]. These results were consistent with previous studies that transcription patterns do not always directly correlate with protein expression levels [16,29,30].
{kind=link}
{kind=link}
{kind=link}