Macrophage activation is involved in laser-induced CNV in a mouse model of AMD
Recent studies have shown that macrophages are recruited to the periphery of the retinal laser photocoagulation induced wet AMD [21], and the activation of M1 and M2 macrophages was examined in the laser-induced CNV mice. Consistent with the previous studies, the CD206+ & F4/80+ macrophages were found to infiltrate the periphery of the laser-injured area in the RPE-choroid in aged mice 7 days post-injury (dpi). At the same time, there was a slight increase in the CD80+ & F4/80+ macrophage levels in the laser-induced area (Figure 1A and 1B). The expression levels of the mRNA for the M2-specific markers including Il-10, arginase-1 (Arg1), mannose receptor C type 2 (Mrc2), Cd301b (Mgl2), and Chil3 (Ym1) or the M1 specific markers, Inos (Nos2), Tnfα, Cd16, and Cd32 also confirmed the M2 and M1 polarization at 7 dpi, respectively (Figure 1C).
To determine whether the M2 macrophages contribute to the development of CNV, the macrophages were depleted by IVI of 2 μl/eye of 90 μg clodronate liposomes as previously described [21]. FFA was performed 14 days after laser photocoagulation to observe vascular leakage and establishment of new vessels. There were several well-defined hyperfluorescent leaking spots representing dye leakage at 14 dpi, while there were fewer leakage spots in those of the clodronate liposomes treatment group (Figure 1D). Macrophage depletion reduced CNV volume at 14 dpi in the clodronate liposomes-treated mice (vehicle control-treated AMD mice, 13.7 × 105 μm3; clodronate treated AMD mice, 2.4 × 105 μm3) (Figure 1E and 1F). Moreover, as shown by OCT, the CNV volume at 14 dpi was also decreased in the clodronate liposomes-treated group compared to the vehicle group (Figure 1G). These results suggest that M2 polarization of macrophages exacerbates laser-induced CNV.
COX2/PGE2 signalling mediates the laser-induced CNV in a mouse model of AMD
PGE2, a metabolite of arachidonic acid catalysed by cyclooxygenase, is involved in the processes of infection, inflammation, immune diseases, and tumour growth [13]. We evaluated the effect of prostanoid-associated signalling in the progression of laser-induced CNV. COX1 showed no significant change in the flat-mounted RPE-choroid at 7 dpi (Figure 2A). COX2 was dramatically increased in the periphery of the laser-injured area at 7 dpi in the RPE-choroid in the aged mice (Figure 2A and 2B). Moreover, as shown by western blot analysis, the amount of COX2 protein at 7 dpi was higher in the laser-treated eyes (Figure 2C and 2D). Consistent with the COX2 inhibitor, clodronate liposomes also significantly decreased the PGE2 production at 7 dpi in the vitreous fluid of the AMD mice (Figure 2E). These data suggest that COX2/PGE2 signalling is critical for the progression of laser-induced CNV.
Celecoxib treatment ameliorates the progression of CNV in a mouse model of AMD
Celecoxib is a highly selective COX2 inhibitor with anti-inflammatory and analgesic properties [22]. In the laser-injured mice, the effects of COX2 on retinal vascular leakage were evaluated. As shown by FFA, there were several intense hyperfluorescent spots at 14 dpi in the laser-injured mice, while there were fewer leakage spots in the celecoxib intravitreal injected (IVI) or celecoxib fed mice (Figure 3A and 3B).
Moreover, the CNV volume shown by OCT and iB4 staining was decreased at 14 dpi in the celecoxib-treated groups, compared to the vehicle control group (average area in the AMD mice, 10.7 × 105 μm3; celecoxib injection mice, 1.85 × 105 μm3; celecoxib fed mice, 1.64 × 105 μm3) (Figure 3C–E). Histological analysis revealed that the laser-induced AMD mice developed robust neovascularization at 14 dpi in the retinal and choroidal tissues, which was partially restored in the celecoxib group (AMD group, 1.25 × 104 μm2; celecoxib feeding group, 0.33 × 104 μm2; celecoxib injection group, 0.23 × 104 μm2) (Figure 3F and 3G).
Celecoxib treatment ameliorates the activation of macrophages in the laser injured mice
To investigate whether the blockade of COX2/PGE2 signalling attenuates macrophage activation, the mice were treated by an IVI of celecoxib, immediately following laser photocoagulation. We observed that the CD206+F4/80+ macrophages infiltrated the periphery of the laser-injured area at 7 dpi in the RPE- The IVI of celecoxib significantly reduced the CD206+F4/80+ macrophages in the RPE-choroid compared to that of the vehicle control. However, the CD80+F4/80+ macrophages infiltrated the periphery of the laser-injured area at 7 dpi in the celecoxib-treated AMD mice showing a slight change vs vehicle control-treated AMD mice (Figure 4A and 4B). The polarization of BMDMs was confirmed by examining the expression levels of mRNA at 7 dpi for the M2-specific markers or M1 specific markers in the laser-induced eyes (Figure 4C and 4D).
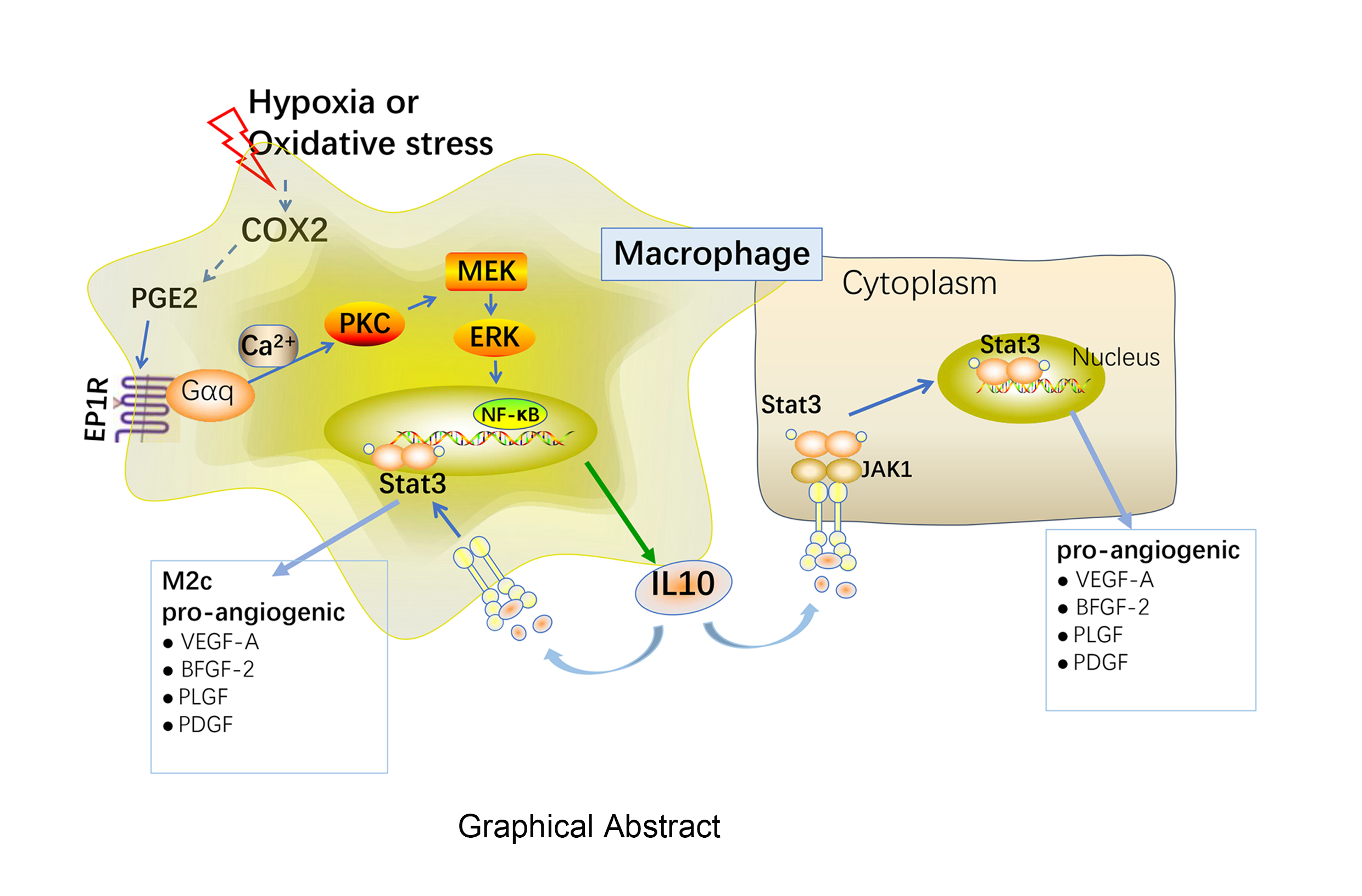
PGE2/EP1R/PKC signalling is involved in the activation of M2 macrophages
PGE2/COX2 signalling was demonstrated to play an important role in M2 macrophage activation and the pathogenesis of the laser-induced CNV. COX2 catalyses the downstream production of prostanoids, including PGE2, which further bind to the EP1–4Rs, inducing the activation of multiple signal transduction pathways that result in the downstream responses [13]. LPS or IFN-γ induce a classical activation of macrophages (M1), whereas IL-4 or IL-13 induce an alternative activation program in the macrophages (M2) [8]. We further examined whether EP1–4Rs are involved in CNV formation in the macrophages in vitro. We isolated the mouse bone marrow cells in vitro by adding M-CSF to induce differentiation into BMDMs after 7 days. Immunofluorescence staining revealed that the positive rate of the F4/80-labeled BMDMs reached 95% (Figure 5A). The M2 polarization of BMDMs significantly induced EP1R protein expression at various time points (24 h and 48 h) in response to the IL-4-stimuli, while the M2 polarization also showed a slight increase in the EP3R expression at 48 h (Figure 5B–F). The BMDMs were treated with PGE2 and an EP1R agonist (17-PT-PGE2), EP2R agonist (Butaprost), EP3R agonist (Sulprostone), and EP4R agonist (Cay10598) to investigate whether EP1-4Rs are the potential regulators of M2 polarization. IL-4 was used as a positive control. Indeed, 5 μM PGE2 or EP1R agonist could induce the activation of M2 macrophages at 24 h (Figure 5G and 5H). As PKA, PKB/Akt, and PKC are the critical intracellular kinases involved in PGE2, activating the EP1-4Rs signalling, the effects of a PKA inhibitor (H89), PKB inhibitor (LY294002), and PKC inhibitor (Bis1) were examined in the PGE2-induced M2 polarization. The Bis1 pre-treatment for 30 min significantly decreased the PGE2-induced M2-specific markers expression at 24 h, while there was no significant difference in the H89 or LY294002-treated groups (Figure 5I). Meanwhile, the EP1R was co-expressed with the laser-induced CNV at 7 dpi in the RPE-choroid mounts of the aged mice (Figure 5J). Taken together, these data suggest that PGE2/EP1R/PKC signalling mediates the activation of the M2 macrophages in the laser-induced CNV.
H2O2 induces the polarization of M2 macrophages of BMDMs
The pathogenesis of wet AMD involves a variety of cellular processes, including oxidative stress, abnormal cell metabolism, and impaired cell function [2]. To test the consequences of increased oxidative stress on macrophage activation of BMDMs, the cells were incubated in different concentrations of H2O2 (0, 100, 200, 400, and 800). After 8 h, there was a significant increase in the expression of the M2-specific marker, while there was no significant difference in M1-specific markers expression between the groups (Figure 6A and 6B). H2O2 treatment for 24 h resulted in a significant increase in the COX2 expression compared to the control medium (Figure 6C and 6D).
PGE2 exacerbates the IL-10 secretion level by increasing the NF-κB binding activity
As an M2-specific marker, IL-10 promotes alternative macrophage activation and pathological neovascularization [10]. In our study, we also found that IL-10 was significantly increased at 7 dpi in the laser-induced eyes, which was reversed by the pre-treatment with celecoxib (Figure 7A). We further identified the region of the IL-10 promoter that was important for mediating the inductive effects of PGE2. The transient transfections were performed with a series of human IL-10 promoter-deletion constructs (Figure 7B). Treatment of the HEK293 cells with PGE2 led to a several-fold increase in the IL-10 promoter activity when the -2000 bp deletion construct (I1) was used (Figure 7C). The magnitude of PGE2-mediated induction of the IL-10 promoter activity remained essentially constant until the -516 bp deletion construct (I3) was used. The -516 bp IL-10 promoter construct (I4) was not stimulated by PGE2. This result implied that one or more promoter elements located between -516 bp and -1012 bp is necessary for the PGE2-mediated induction of the IL-10 promoter activity. NF-κB sites are found within this region of the IL-10 promoter [23]. Recent studies have reported that NF-κB plays important role in the early stages of angiogenesis and CNV [24]. NF-κB is also known to regulate several genes involved in angiogenesis (VEGF, intercellular adhesion molecule 1 [ICAM1] and COX2) [25, 26]. As oxidative stress serves a vital function in the pathogenesis of AMD, in our study, the BMDMs were also subjected to H2O2 treatment to induce oxidative damage. The results revealed that 400 μM H2O2 or 5 μM PGE2 dramatically induced the DNA binding activity of NF-κB (Figure 7D). Meanwhile, treatment of the BMDMs with H2O2 or PGE2 could increase the NF-κB translocation from the cytoplasm to the nucleus (Figure 7E and 7F). Besides, 10 μM celecoxib treatment dramatically prevented the H2O2-induced p65 expression in the nucleus (Figure 7G and 7H).
MAPKs, including Erk (1/2), p38 and JNK (1/2), mediated the activation of NF-κB, which have been demonstrated to play an important role in oxidative stress [27]. As shown in Figure 7, H2O2 induced a significant increase in the phosphorylation of Erk (1/2) compared to the control group, which was prevented in the celecoxib-treated group (Figure 7I and 7J). However, there was no significant difference in the level of phospho-p38 or phospho-JNK (1/2) among the groups (Figure 7I and 7J).
Role of IL-10 in the proliferation and migration of the human choroidal microvascular endothelial cells (HCECs)
The role of IL-10 in HCECs proliferation and migration were examined using the WST-1 and transwell migration assays, as well as by observing the distance in wound healing. The proliferation and migration were dramatically increased in the 10 ng/ml or 25 ng/ml IL-10-treated HCECs (Figure 8A–8C).
{kind=link}