Virotherapy with GLV-1h68
First, effects of a monotherapy of the six NET/NEC cell lines employing the vaccinia virus vector GLV-1h68 were studied. In this purpose, SRB viability assays were conducted to evaluate cytostatic and cytotoxic effects of the OV on neuroendocrine cancer cells and to identify oncolysis-sensitive and -resistant tumor cell lines. Further, microscopic fluorescence pictures were taken to visualize oncolysis and directly detect and prove virotherapeutic vector-based transgene (GFP) expression. Next, a real-time cell monitoring assay was employed to distinguish between cytostatic and cytotoxic nature of the effect and study the dose dependency of this circumstance. Finally, the production of viral progeny, which forms the basis of the intratumoral infectious spread of an OV, was studied by assessing virus titers sequentially over time.
Oncolysis with GLV-1h68
All NET/NEC cell lines were infected with multiplicities of infection (MOIs) of GLV-1h68 in logarithmic steps, ranging from 0.0001 to 1. Taking the first results of the SRB viability assays into account, the MOIs were modified by adding MOI 0.5 instead of MOI 0.0001 for all cell lines except BON-1; MOIs 0.025 and 0.05 were added for BON-1 cells, while MOIs 0.0001 and 0.001 were left out. A threshold for clinically relevant anti-tumor activities was set at 60 % of tumor cells being residual in SRB viability assays after an infection period of 96 hours (Figure 1, dotted horizontal lines). Three categories to classify cellular response to GLV-1h68 virotherapy were introduced: (i) highly permissive cell lines, meeting the 60 % threshold with MOI 0.1 or less after 96 hours; (ii) permissive cell lines requiring MOI 0.5 to meet the threshold at 96 hpi, and (iii) resistant cell lines which required more than MOI 0.5 to meet the threshold at 96 hpi.
It was found that GLV-1h68 is able to infect and kill all six NET/NEC cell lines, requiring different MOIs for the same effect. For all tumor cell lines, a dose dependency was observed, meaning that a higher MOI resulted in a lower number of residual tumor cells at the end of the observation period, i.e. at 96 hpi. As a result, two highly permissive, three permissive and one resistant cell lines could be identified. BON-1 pancreatic NET (pNET) cells were found to be most sensitive to GLV-1h68-mediated oncolysis, exhibiting a remaining tumor cell mass of 60 % at 96 hpi when using a MOI of only 0.01 (Figure 1C). For all other NET/NEC cell lines higher MOIs had to be applied in order to meet the 60 % threshold at 96 hpi: MOI 0.1 was enough for HROC-57 cells (Figure 1E); accordingly, BON-1 and HROC-57 cells were classified as highly permissive. In contrast, H727, QGP-1, and NEC-DUE1 cells required MOI 0.5 and were classified as permissive. UMC-11 cells were found to be the only resistant cell line to GLV-1h68-mediated cell killing, closely reaching 60 % remnant tumor cell mass with MOI 1 at 96 hpi (Figure 1B).
Overall, GLV-1h68 was able to reduce the tumor cell masses to a minimum of less than 10 % in 4 out of 6 NET/NEC cell lines. Regarding the results of this experiment, higher, but impractical MOIs potentially would be able to further reduce the remnant tumor cell count even in the oncolysis-resistant UMC-11 cell line.
In summary, only a single neuroendocrine cancer cell line turned out to be resistant to GLV-1h68-mediated oncolysis. The two highly permissive cell lines were found to be BON-1 originating from a pNET and HROC-57 originating from a colon NEC, whereas the lung NET derived cell line UMC-11 was shown to be the most resistant cell line. Given that the three other cell lines showed very similar responses, no obvious relation between anatomical origin and treatment response could be identified in this experiment.
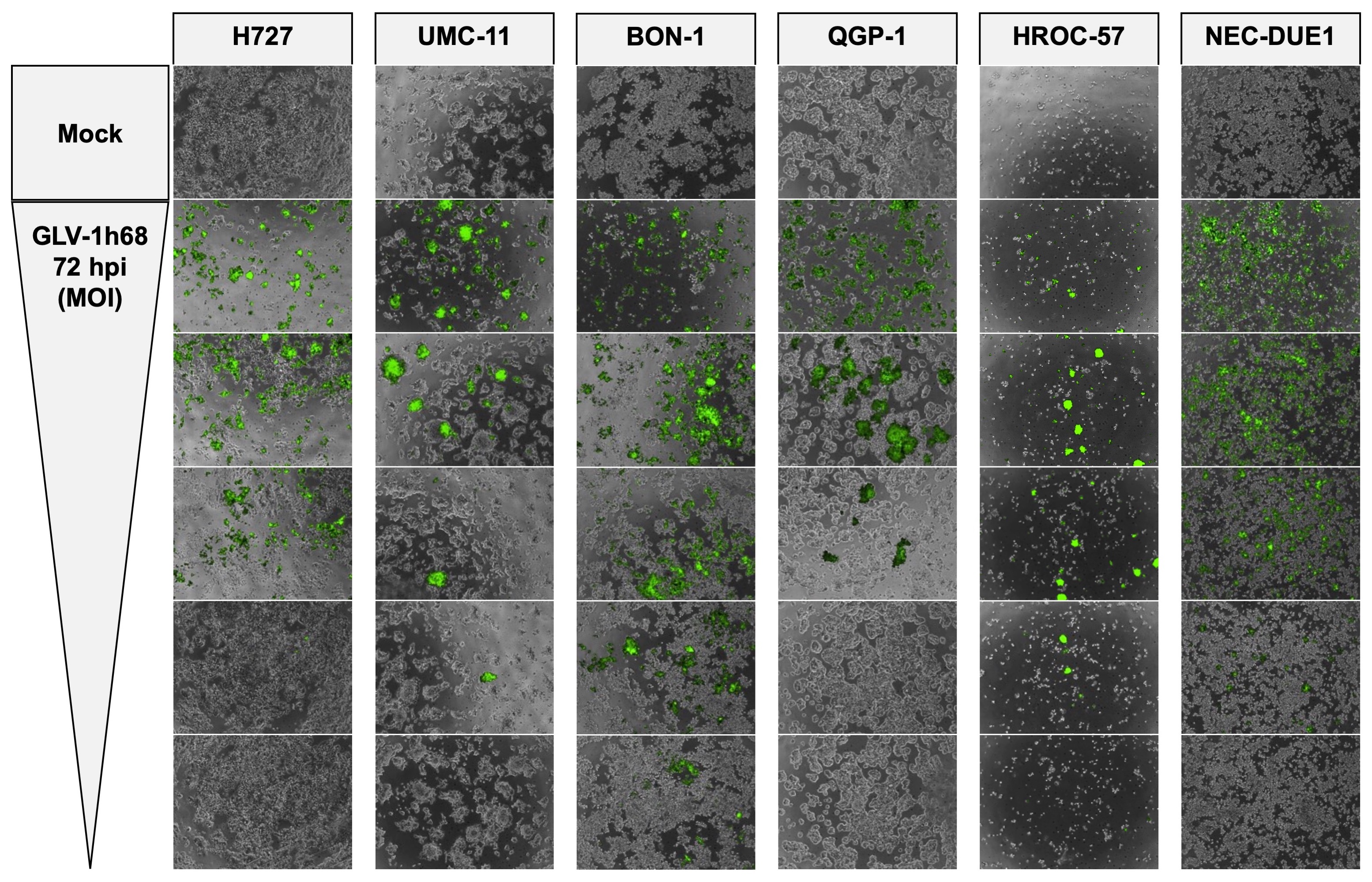
Microscopy of GLV-1h68-mediated NET/NEC cell oncolysis
As GLV-1h68 encodes a fluorescent GFP transgene for therapeutic monitoring, microscopic pictures were taken to prove viral infection and replication via transgene expression and observation of cell layer densities (Figures 2 and S1). The same MOIs as in the SRB viability assay (Figure 1) were applied. As a result, a loss of cell density could be observed in all infected neuroendocrine cancer cell lines, consistent with results from the SRB viability assay, where all tumor cell lines were found to respond to virus infections. Moreover, all analyzed NET/NEC cell lines were found to express the GFP transgene when being infected with GLV-1h68. Of note, lower cell confluency and intensities of the fluorescence signals were found to correlate to the MOIs being applied (Figures 2 and S1). This does not apply for HROC-57 cells, as the confluency was also low in uninfected cells (mock). However, with the highly permissive cell lines (BON-1, HROC-57), the highest MOI displayed lower transgene expression, most likely because of a high rate of oncolysis and therefore a lower cell count expressing the fluorescent GFP transgene. This phenomenon is also visible with permissive QGP-1 cells and on the respective pictures taken at 72 hpi, although to a lesser extent (Figure S1). Mock treatment did not display any fluorescence at all.
Real-time cell monitoring
To precisely investigate the nature of the effect of GLV-1h68 on neuroendocrine cancer cells, a real-time cell monitoring assay was employed. The lung NET cell line H727 was picked as representative cell line because it showed a stable, average response to GLV-1h68 in the experiments described above. Two MOIs (0.1 and 0.25), which resulted in remaining tumor cell numbers of around 50 % according to SRB viability assay performed at 96 hpi, were chosen for infection. The xCELLigence RTCA assay measures cellular impedance, which was shown to correlate with cell number, cell size/morphology and cell attachment quality [35]. Taking the previous SRB viability assays and applied cell lysis control with Triton X-100 into account, the Cell Index can be seen as a surrogate for cell viability in this context.
Different treatment modalities were initiated at 24 hours after cell seeding. As expected, treatment with the cell lysis control Triton X-100 immediately resulted in a complete tumor cell lysis (Figure 3; green dotted line). In contrast, virus infections showed similar results to mock treatment in the first 24 hpi. In the further course of the experiment, the impedance of infected cells decreased continuously, indicating not only a cytostatic but also a cytotoxic effect of GLV-1h68. The higher MOI (0.25) results in lower cell viability in the end, but not in a faster mechanism of action, also showing the first impairment of tumor cell growth at 24 hpi and the peak of cell viability at 36 hpi (Figure 3; line with grey squares). Taken together, GLV-1h68 was proven to exhibit a pronounced oncolytic effect on the neuroendocrine tumor cell line H727 and also a dependency on the infectious dose being applied. Thus, findings of the SRB viability assay could be confirmed.
Virus titer quantification
As the production of viral progeny is an important step in the underlying mechanism of oncolytic virotherapy, virus titers obtained by neuroendocrine cancer host cells were sequentially determined every 24 hours during the whole period of infection. Hence, the lung NET cell line H727 (Figure 4A) and the pNET cell line BON-1 (Figure 4B), which was found to be the tumor cell line being most sensitive to GLV-1h68 treatment, were picked to further investigate tumor cells being established from different anatomical origins. Both NET cell lines were infected with MOIs achieving around 50 % reductions of tumor cell counts in the SRB viability assays. Shortly after virus infection, all extracellular viral particles were removed so that only viral particles which had already entered the cells after a 1-hour infection period could produce viral progeny.
As a result, high levels of viral replication could be detected in both tumor cell lines (Figure 4). Titers over 107 plaque forming units (PFU)/ml were easily reached within 72 hours. A stagnation of virus titer growth could be observed for H727 after 72 hours and a reduction of viral titer between 72 and 96 hours was detected with BON-1 cells.
Combinatorial treatment with everolimus
Next, a combinatorial treatment with the mTOR inhibitor everolimus was evaluated by comparing a combinatorial approach (GLV-1h68 + everolimus) to GLV-1h68 monotherapy. In this purpose, SRB viability assay and virus quantification were conducted.
Oncolysis with GLV-1h68 and everolimus
SRB viability assays were carried out using the lung NET cell line H727 and the NEC cell line NEC-DUE1, which both are tumor cell lines being generated from different anatomical origins, but both were classified as permissive to GLV-1h68 monotherapy (Figure 1). Again, MOIs leading to around 50 % tumor cell reductions were chosen (0.1 and 0.25 for H727; 0.25 and 0.5 for NEC-DUE1). Everolimus was administered in concentrations of 1 nM for H727 cells and 0.25 nM for NEC-DUE1 cells, respectively.
As a result, the addition of GLV-1h68 to sole everolimus treatment was found to be able to further reduce the remaining tumor cell count. This was observed in both cell lines tested and with both MOIs employed in each cell line. Interestingly, the benefit of the combinatorial therapy appeared to be more pronounced in NEC-DUE1 cells. However, the extent of this effect was limited in its extent, thereby not representing any additive mechanism of action.
Virus titer quantification
To investigate whether everolimus has any impact on virus replication, virus titers were assessed when GLV-1h68 was employed in a combinatorial setting with everolimus (Figure 6, dotted lines). In both NET cell lines (H727, BON-1), where virus replication was determined previously, everolimus did not affect the production of viral progeny in any way.
Taking the results from both assays into account, the final benefit of the combinatorial therapy after 96 hours is visible but only small. Everolimus did not limit virus replication in a particular way. Given that evidence base, the combinatorial therapy of GLV-1h68 with everolimus was not found to be inferior to either monotherapy and can be regarded as a possible future combinatorial treatment option for metastatic neuroendocrine cancer.
{kind=link}