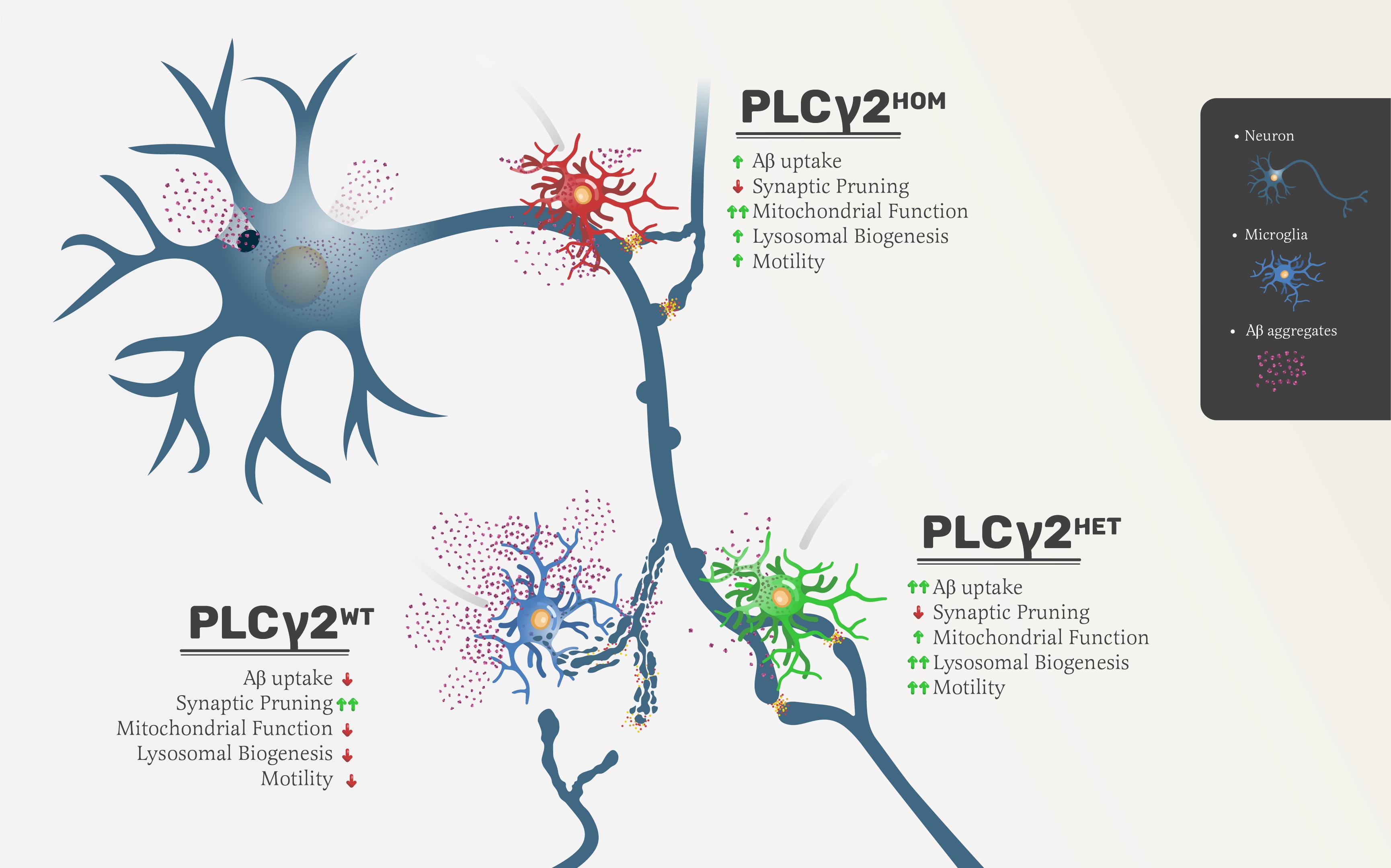
In this study, we investigated the impact of the AD protective PLCγ2P522R variant on muman microglial cell functions. We used CRISPR editing to generate PLCγ2HET and PLCγ2HOM hiPSC cells, and differentiated them into microglia that exhibit functional and transcriptional profiles consistent with human microglia [13]. We were able to show that the presence of the P522R variant resulted in increases in PLCγ2 levels, as previously reported [8]. It also increased microglial uptake of the Aβ peptide, accompanied by a reduction in synapse pruning, two key pathological processes in AD. We provide evidence that this discrimination appears to be primarily driven by cargo size, and that cells expressing the PLCγ2P522R variant have enhanced mitochondrial function and cell motility, potentially associated with increased intracellular Ca2+. We have also identified transcriptional changes in a number of genes associated with lipid metabolism, endosome/phagosome maturation and immune function in resting state PLCγ2HET microglia, which highlight potential pathways that may underpin the protective capacity of the PLCγ2P522R variant in AD. Crucially, we have shown that the impact of PLCγ2P522R is dependent on dose, with heterozygous cells overall showing a more beneficial profile than homozygous cells.
The clearance of Aβ, along with other cellular debris, is an important feature of microglial phagocytic function in AD. Several other polymorphisms implicated as risk factors in AD, such as the R47H TREM2 mutation, result in alterations to microglial phagocytic capacity that lead to reduced clearance of Aβ aggregates and neuronal decline [18, 19]. In our hiPSC derived microglia assay, PLCγ2P522R enhanced the uptake of Aβ compared to PLCγ2WT microglia, suggesting that the variant may positively impact directly on disease relevant pathology clearance, as previously suggested [12]. Consistent with this increased Aβ clearance, lysotracker levels, which indicate acidic vesicles such as lysosomes, were higher in both PLCγ2HET and PLCγ2HOM microglia variants compared to PLCγ2WT cells. Interestingly, PLCγ2HET microglia showed a more robust increase in lysotracker levels than PLCγ2HOM cells, suggesting that there may be a narrow functional level (“sweet spot”) of PLCγ2P522R that leads to improved outcomes. This is supported by the fact that while a number of the processes we assessed did appear to be ‘dose dependently’ modified by heterozygous vs. homozygous expression of the PLCγ2P522R variant, other processes were preferentially impacted only in heterozygous cells. Indeed, a number of gain of function mutations in PLCγ2 have been linked to immune disorders, including PLAID (PLCγ2-associated antibody deficiency and immune dysregulation syndrome), APLAID (autoinflammation, antibody deficiency and immune dysregulation syndrome) and FCAS3 (familial cold autoinflammatory syndrome), as well as a subset of CVID (common variable immunodeficiency) patients [20–27]. These clearly indicate that a significant gain of function of PLCγ2 can lead to deleterious impacts on the immune system.
In the central nervous system, microglia shape neuronal synaptic connections and strength during development, through the removal of excess synapses [28]. While microglia mediated synaptic pruning is a fundamental physiological process during development, its proposed reactivation during aging has deleterious consequences and may account for much of the memory loss and cognitive decline observed in AD [15]. It has recently been reported that individuals with MCI (mild cognitive impairment), who carry the PLCγ2P522R variant had better cognitive performance even in the presence of the APOE4 AD risk gene [29]. Here we show hiPSC microglia with the PLCγ2P522R variant had decreased synaptosome uptake compared to WT cells. Moreover, we were able to demonstrate that these PLCγ2P522R microglia selectively spare synaptosomes while maintaining efficient clearance of Aβ when treated concurrently with both biological cargoes. These finding were replicated in BV2 cells over expressing exogenous human PLCγ2WT or PLCγ2P522R, indicating a possible connection between increased expression of PLCγ2 and reduced synapse uptake. We provide further evidence of this in our neuronal and microglial co-culture study, where we saw a reduction in synaptic pruning by PLCγ2P522R microglia, as evidenced by reduced levels of PSD95 within microglial cells. Interestingly, this was once again more robustly observed in PLCγ2HET cells, supporting the notion of a ‘sweet spot’ for maximal beneficial impacts on microglial function.
The most prevailing question is how does the PLCγ2P522R variant modulate diverse microglial functions to protect a degenerating brain? Our gene expression study highlights a number of different functional pathways that appear to be activated in PLCγ2HET microglia, notably those relating to lipid metabolism, endosome/phagosome maturation and immune function. Proinflammatory environments and signalling are a prominent observation in AD brain pathology [30]. Our findings suggest a significant increase in the expression of the anti-inflammatory cytokine, interleukin-10 (IL-10), which is known to inhibit the synthesis of pro-inflammatory cytokines, and may promote the differentiation of microglia into the anti-inflammatory state, classically known as M2 or homeostatic state [31]. Interestingly, IL-10 deficiency has also been shown to exacerbate Tau pathology [32]. This, combined with our data, provides a potential route whereby the PLCγ2P522R variant may impact at least in part on both primary pathologies associated with AD to reduce disease risk.
We also report significant increases in the expression of the chemokine receptor, CX3CR1 in PLCG2HET cells. Within the CNS, this receptor is expressed exclusively by microglia, and binds to the neuronally expressed chemokine, CX3CL1 (fractalkine). The exact function of this interaction is not yet clear, but evidence suggests it plays an important role in the regulation of synaptic function, with CX3CR1 deficiency resulting in cognitive and LTP impairments indicative of altered synaptic plasticity [33, 34]. Certainly increased expression of CX3CR1 attenuates microglial inflammatory responses to LPS (liposccharide) (Inoue et al., 2021, PLOS One), while deletion results in an increased inflammatory response and phagocytic acivity (Murai et al, 2020, Eur J Neurosci). It is therefore possible that the increase in CX3CR1 we observe in resting state PLCγ2HET microglia is linked to the reduction in synapse phagocytosis seen in these cells, although more work is required to unpick the mechanism that may underpin this.
A growing body of evidence suggests that lipid metabolism is crucial to fuel microglial functions such as phagocytosis, and can vary depending upon the activation status of the cell (reviewd in [35]). Previous studies have shown that PLCG2 acts downstream of TREM2, and knock out of either protein results in lipid accumulation deficits and a failure to activate key lipid processing genes such as LIPA and APOC1 [11], as well as a generalised alteration in the overall lipidome of the cells. In the current study, heterozygous PLCγ2P522R induced increases in the expression of genes associated with phagocyte maturation (RAB5, RAB7) and various aspects of lipid metabolism (LIPA, APOE, PLIN2) in resting state microglia, an effect which is not seen in homozygous cells. This may suggest that these heterozygous cells are ‘primed’ to more readily respond to an inflammatory challenge, and can therefore more efficiently uptake and phagocytose targets such as Aβ when required.
It is important to acknowledge that the gene expression changes we observed in the PLCγ2HET microglia were all seen in resting state cells, without LPS or other disease relevant immune challenge. These expression changes are likely to be important factors in the protective capacity of PLCγ2P522R. LOAD develops over many years, rather than arising as a result of a single acute challenge, thus these low level ongoing changes likely reflect the chronic impact of the variant on general microglial function. However, it is probable that additional changes will be observed in response to immune challenge, and further work is required to eludicate these changes, and understand how they impact on microglial function, to improve target cargo uptake and preserve synapses. Indeed, we identified a number of additional genes that trended towards a difference between PLCγ2HET and PLCγ2WT cells, that may indicate a subset of genes ‘primed’ to respond to immune challenge. Loss of PLCG2 results in a spectrum of gene expression changes in microglia both at rest, and in response to TLR stimulation [11], and intriguingly, this TLR induced profile appears distinctly different from that seen with TREM2 knockout, suggesting that PLCG2 may interact independently with both TREM2 and TLR linked pathways, although we did not see significant changes in the basal expression of TREM2 or TLR4, or functionally associated proteins such as NLRP3 in our cells. A broad spectrum of genes expression changes have also been reported in homozygous PLCγ2P522R knock-in mice [36]. However, given our findings, which suggest that heterozgosity is important for maximimal benefits from the variant, examining the difference between heterozygous and homozygous PLCγ2P522R basal and stimulated gene expression profiles may be invaluable in more clearly identifying specific functional changes responsible for the protective impact of PLCγ2P522R in LOAD.
The highly dynamic nature of microglia makes them heavily dependent on efficient energy expenditure to meet basic housekeeping demands. To maintain brain homeostasis and surveillance, microglia require energy-demanding cytoskeleton remodelling, necessary for extending and retracting their ramified processes, to constantly scan their surroundings [37, 38]. Whilst in a resting state, microglia use mitochondrial respiration, specifically OXPHOS (oxidative phosphorylation) as their main source of energy, although resting state is a misconception, since microglia are never quiescent, rather they are constantly engaged in parenchymal surveillance [38]. Our findings indicate that microglia expressing PLCγ2P522R in general show enhanced mitochondrial respiration as well as significantly higher ATP production, which was not associated with increases in mitochondrial number but rather changes in mitochondrial function.
Mitochondrial function directly correlates with cytoplasmic Ca2+ levels, which could underlie the enhancement we observed in the PLCγ2P522R cells. PLCγ2 is a direct modulator of Ca2+ influx from the endoplasmic reticulum (ER) via its hydrolysis of PIP2, leading to the production of IP3, which then interacts with its receptor IP3R1 [39]. Recent reports have demonstrated that the presence of the PLCγ2P522R variant in microglia enhances Ca2+ release in response to physiological stimuli [12]. In our study, we found that PLCγ2HET microglia showed higher levels of basal intracellular Ca2+ than PLCγ2WT cells in the absence of any stimulus, supporting the notion that the PLCγ2HET cells may be ‘primed’ to respond more rapidly to an inflammatory challenge. This increase was not observed in the PLCγ2HOM cells, once again highlighting the delicate balance of P522R dose on PLCγ2 function. Interestingly, PLCγ2HOM cells did demonstrate a greater enhancement of mitochondrial function than the PLCγ2HET cells, in the absence of any change in Ca2+ levels. This separation between Ca2+ levels and mitochondrial performance suggests that other as yet unidentified factors may play a key role in modulating microglial cell energy homeostasis.
Ca2+ currents have also been implicated in the regulation of microglial motility (reviewed in [40]. Consistent with the increase we see in intracellular Ca2+ in PLCγ2HET cells, they also demonstrated enhanced mobility compared to PLCγ2WT cells. Both whole cell movement, detected via nuclear tracking, as well as surveillance, detected via assessing cytoplasmic movement alone, were enhanced. PLCγ2HOM cells also showed increased surveillance, but their nuclear movement did not differ from PLCγ2WT cells, once again highlighting the delicate functional balance between heterozygous and homozygous expression of the P522R variant.
Our study is the first to suggest that the protective nature of the PLCγ2P522R variant in AD may reflect a selective increase in microglial clearance of Aβ, coupled with a preservation of synpases. This is accompanied by an increase in mitochondrial fitness and microglial motility, suggesting that PLCγ2P522R may enhance basal microglial functioning, leading to a relative protection against the accumulation of pathogenic proteins seen in both healthy aging and AD. A number of resting state transcriptional changes are associated with this altered function, which appears to be critically dependent on PLCγ2P522R ‘dose’, with heterozygous expression of the variant optimal.
{kind=link}